Schema pentru obținerea hibrizilor din prima generație. Prima lege a lui Mendel este legea dominației. Legea purității gametilor și raționamentul său citologic
6. Ce este ontogenia. Care sunt perioadele de ontogeneză? Faceți o scurtă descriere a dezvoltării postembrionare.
1. Nivelurile organizațiilor faunei sălbatice (listă).
2. Numiți organele celulare cu două membrane și non-membrane. Indicați caracteristicile structurii și funcției lor.
3. Care sunt avantajele procesului de reproducere sexuală față de asexual.
4. A treia lege a lui Mendel. Indicați raportul descendenților după genotip și fenotip.
5. Numiți centrele de origine și diversitatea plantelor cultivate conform lui Vavilov. Unde este stocată colecția de semințe de plante care crește pe Pământ, care este semnificația acesteia.
6. Ce este ontogenia. Care sunt perioadele de ontogeneză? Faceți o scurtă descriere a dezvoltării postembrionare.
corpul gazdei. Măsuri preventive.
2. Forțele motoare ale evoluției și relația lor.
5. Determinați mărirea microscopului. Pregătește-l pentru muncă.
Numărul biletului 6
1. Sistemul circulator al vertebratelor.
2.1. Traversare monohibridă.
2. Prima și a doua lege a lui Mendel.
6. Folosind material de ierbariu, grupați plantele în familii.
Numărul biletului 7
1. Departamente ale sistemului nervos: central și periferic. 2. Habitatul organismelor.
Biologie, ajutor !!!1. Știință care studiază tiparele de transmitere a trăsăturilor prin moștenire
2. Au fost formulate mai întâi tiparele genetice
3. Cromozomi împerecheați, fiecare dintre aceștia mergând în corp de la unul dintre părinți
4. Cromozomi care conțin un set de gene identice
5. Cromozomi localizați în același organism, dar diferiți ca mărime, formă și set de gene
6. Gene situate în aceleași regiuni ale cromozomilor omologi, responsabile de sinteza acelorași proteine, dar având o secvență nucleotidică diferită
7. Care va fi numele genelor care codifică aceeași proteină (de exemplu, hemoglobină sau melanină), dar care conțin o secvență de aminoacizi diferită
8. La traversarea monohibridă, semnele sunt investigate
9. Conform genei culorii (în experimentele lui Mendel), mazărea avea numărul de trăsături
10. zigot care a primit două alele diferite
11. zigot care a primit ambele aceleași alele
12. Gena notată când este traversată de o literă mică
13. Care este numele alelei, notată la traversarea unei litere majuscule (majuscule)
14. O alelă care apare fenotipic doar într-o stare homozigotă
15. Alela, care se manifestă extern în orice caz, dacă este prezentă deloc în genotipul unui individ
16. Genotipul zigotului denumit AA
17. Genotipul zigotului denumit Aa
18. Genotipul zigotului, desemnat aa
19. Ansamblul semnelor externe ale corpului
20. Totalitatea tuturor caracteristicilor ereditare ale organismului
21. Ansamblul tuturor genelor unei specii sau ale unei populații
22. Împărțirea hibrizilor în a doua generație urmează formula 3: 1
23. Genotipul formelor parentale de mazăre („linii curate”) în experimentele lui Mendel
24. Genotipul descendenților (hibrizi din prima generație) în mazăre în experimentele lui Mendel
25. Culoarea mazărei de prima generație
26. F1 Mendel a obținut prin polenizare
27. F2 Mendel obținut prin polenizare
28. Tipul de clivaj la traversarea heterozigotului și homozigotului pentru recesivitate
29. Formele monoheterozigote ... soiuri de gameti
30. Hibrizi cu încrucișare monohibridă și formă de dominanță completă ... clase fenotipice
31. Legile lui Mendel se refereau la acest tip de moștenire
32. Genele non-alelice situate pe același cromozom, dacă nu are loc încrucișarea, sunt moștenite
33. Autorul teoriei cromozomiale a eredității
34. Cromozomi care diferă ca mărime și formă la bărbați și femei
35. Cromozomi care nu diferă ca mărime și formă
36. Genele „legate de sex” sunt localizate
37. Hemofilia și orbirea culorii la oameni și moștenirea culorii hainei la pisici sunt exemple de
38. Genele acestor trăsături sunt localizate
39. Mamiferele masculine și Drosophila sunt heterozigoți pentru trăsături legate de sex
40. Modificări accidentale ale materialului genetic al unui individ
41. Modificarea numărului de cromozomi
42. Schimbarea unuia sau mai multor nucleotide
43. Modificări ale secvenței de aminoacizi a proteinelor
44. Autorul legii seriilor omoloage ale variației ereditare
45. Principala metodă de genetică, inaplicabilă în genetică umană
46. Metoda geneticii umane, bazată pe colorarea cromozomilor și studiul ulterior al mărimii și formei acestora
47. Metoda geneticii umane, bazată pe analiza genealogiilor și trasarea transmiterii unei anumite trăsături
48. Metoda geneticii umane, bazată pe analiza manifestărilor fenotipice ale trăsăturilor la gemeni identici
49. Metoda geneticii umane, bazată pe analiza activității enzimatice a proteinelor care catalizează procese fiziologice importante
50. Substanțe care cauzează mutații
Introducere.
Genetica este o știință care studiază legile eredității și variabilității organismelor vii.
Omul a remarcat de mult trei fenomene legate de ereditate: în primul rând, asemănarea trăsăturilor descendenților și părinților; în al doilea rând, diferențele dintre unele (uneori multe) trăsături ale descendenților față de trăsăturile parentale corespunzătoare; în al treilea rând, apariția în urmași a unor personaje care erau doar în strămoșii îndepărtați. Continuitatea trăsăturilor între generații este asigurată de procesul de fertilizare. Din timpuri imemoriale, omul a folosit spontan proprietățile eredității în scopuri practice - pentru a reproduce soiuri de plante cultivate și rase de animale domestice.
Primele idei despre mecanismul eredității au fost exprimate de oamenii de știință greci antici Democrit, Hipocrate, Platon, Aristotel. Autorul primei teorii științifice a evoluției J.-B. Lamarck a folosit ideile oamenilor de știință greci antici pentru a explica ceea ce a postulat la începutul secolelor XVIII-XIX. principiul transferului de noi trăsături dobândite în timpul vieții unui individ către descendenți. C. Darwin a prezentat teoria pangenezei, explicând moștenirea personajelor dobândite
A definit Charles Darwin ereditate ca proprietate a tuturor organismelor vii de a le transmite caracteristicile și proprietățile din generație în generație și variabilitate ca proprietate a tuturor organismelor vii de a dobândi noi caracteristici în procesul dezvoltării individuale.
Trăsăturile sunt moștenite prin reproducere. Odată cu reproducerea sexuală, noile generații apar ca urmare a fertilizării. Fundamentele materiale ale eredității sunt conținute în celulele germinale. Cu reproducere asexuată sau vegetativă, o nouă generație se dezvoltă fie din spori unicelulari, fie din formațiuni multicelulare. Și cu aceste forme de reproducere, legătura dintre generații se realizează prin intermediul celulelor, care conțin bazele materiale ale eredității (unități elementare ale eredității) - gene - sunt secțiuni ale ADN-ului cromozomilor.
Setul de gene pe care un organism le primește de la părinții săi constituie genotipul său. Totalitatea trăsăturilor externe și interne este un fenotip. Fenotipul se dezvoltă ca urmare a interacțiunii genotipului și a condițiilor de mediu. Într-un fel sau altul, baza este trăsăturile care poartă genele.
Modelele prin care semnele sunt transmise din generație în generație au fost descoperite pentru prima dată de marele om de știință ceh Gregor Mendel. El a descoperit și a formulat trei legi ale moștenirii care au stat la baza geneticii moderne.
Viața și cercetările științifice ale lui Gregor Johann Mendel.
Călugăr moravian și genetician al plantelor. Johann Mendel s-a născut în 1822 în orașul Heinzendorf (acum Gincice din Republica Cehă), unde tatăl său deținea un mic lot de țărani. Gregor Mendel, conform mărturiei celor care l-au cunoscut, era într-adevăr o persoană amabilă și plăcută. După ce a primit învățământul primar la școala locală din sat și mai târziu, după ce a absolvit Colegiul Piaristilor din Leipnick, a fost admis la Liceul Imperial-Regal Troppaun în 1834 la prima clasă de gramatică. Patru ani mai târziu, părinții lui Johann, ca urmare a confluenței multor evenimente nefericite care s-au succedat rapid, au fost complet lipsiți de ocazia de a rambursa cheltuielile necesare asociate cu educația, iar fiul lor, având atunci doar 16 ani, avea obligat să se îngrijească complet independent de propria întreținere ... În 1843, Mendel a fost admis la mănăstirea augustiniană Sf. Toma din Altbrunn, unde a luat numele de Gregor. În 1846, Mendel a participat și la prelegeri despre agricultură, horticultură și viticultură la Institutul Filozofic din Brunn. În 1848, după finalizarea unui curs de teologie, Mendel a primit, cu profund respect, permisiunea de a studia pentru examenele de doctorat. Când anul următor și-a întărit intenția de a fi examinat, i s-a dat ordin să ia locul susținătorului gimnaziului imperial-regal din Znaim, pe care l-a urmat cu bucurie.
În 1851, starețul mănăstirii l-a trimis pe Mendel să studieze la Universitatea din Viena, unde, printre altele, a studiat botanica. După absolvirea universității, Mendel a predat științe la o școală locală. Datorită acestui pas, situația sa financiară sa schimbat radical. În mult necesitatea bunăstării benefice a existenței fizice pentru fiecare căutare, curajul și puterea i-au revenit cu o profundă venerație și, în timpul anului de încercare, a studiat cu mare sârguință și dragoste subiectele clasice prescrise. În orele libere a fost angajat într-o mică colecție botanică și mineralogică, care se afla la dispoziția sa în mănăstire. Pasiunea sa pentru domeniul științelor naturii a devenit cu atât mai mare, cu cât avea mai multe oportunități de a se preda lui. Deși persoana menționată în aceste lecții a fost lipsită de orice îndrumare, iar calea autodidactului aici, ca în nicio altă știință, este dificilă și duce la obiectiv încet, totuși, în acest timp, Mendel a dobândit o astfel de dragoste pentru studiul naturii, încât nu și-a cruțat eforturile.să-și umple golurile schimbate prin auto-studiu și urmând sfaturile oamenilor cu experiență practică. La 3 aprilie 1851, „corpul didactic” al școlii a decis să îl invite pe domnul Gregor Mendel, canonul mănăstirii St. Succesele pomologice ale lui Gregor Mendel i-au adus dreptul la un titlu stelar și o poziție temporară ca susținător al istoriei naturale în clasa pregătitoare a Școlii Tehnice. În primul semestru al studiilor, a studiat doar zece ore pe săptămână și numai cu Doppler. În al doilea semestru, studia douăzeci de ore pe săptămână. Dintre aceștia, zece - în fizică cu Doppler, cinci pe săptămână - în zoologie cu Rudolf Kner. Unsprezece ore pe săptămână - botanică cu profesorul Fenzl: pe lângă prelegeri despre morfologie și taxonomie, a susținut și un atelier special despre descrierea și definirea plantelor. În al treilea semestru, s-a înscris pentru treizeci și două de ore de curs pe săptămână: zece ore pentru fizică la Doppler, zece pentru chimie la Rottenbacher: chimie generală, chimie medicinală, chimie farmacologică și un atelier de chimie analitică. Cinci pentru zoologia lui Kner. Șase ore de antrenament cu Unger, unul dintre primii citologi din lume. În laboratoarele sale, a studiat anatomia și fiziologia plantelor și a urmat un atelier de tehnici de microscopie. Și din nou - o dată pe săptămână la Departamentul de Matematică - un atelier despre logaritm și trigonometrie.
1850, viața mergea bine. Mendel se putea întreține deja și se bucura de un respect deosebit din partea colegilor săi, pentru că se descurca bine cu îndatoririle sale și era foarte plăcut să vorbească. Ucenicii Lui l-au iubit.
În 1851, Gregor Mendel s-a aplecat asupra problemei cardinale a biologiei - problema variabilității și eredității. Atunci a început să efectueze experimente privind cultivarea dirijată a plantelor. Mendel a adus diverse plante de departe și din apropierea Brunn. El a cultivat plante în grupuri într-o parte a grădinii mănăstirii special concepute pentru fiecare dintre ele în diferite condiții externe. Era angajat în observații meteorologice minuțioase. Gregor a condus majoritatea experimentelor și observațiilor sale cu mazăre, pe care, din 1854, le-a semănat în fiecare primăvară în fiecare primăvară într-o mică grădină sub ferestrele prelaturii. S-a dovedit a fi ușor să organizezi un experiment clar de hibridizare pe mazăre. Pentru a face acest lucru, trebuie doar să deschideți o floare mare, deși nu încă coaptă, cu pensete, să tăiați anterele și să determinați în mod independent „perechea” pentru încrucișare. Deoarece auto-polenizarea este exclusă, soiurile de mazăre sunt, de regulă, „linii curate” cu caracteristici constante care nu se schimbă de la generație la generație, care sunt foarte clar conturate. Mendel a evidențiat trăsăturile care au determinat diferențele intervarietale: culoarea coajei boabelor mature și - separat - a boabelor necoapte, forma mazărei mature, culoarea „proteinei” (endosperm), lungimea axei tulpinii , locația și culoarea mugurilor. El a folosit mai mult de treizeci de soiuri în experiment și fiecare dintre soiuri a fost anterior supusă unui test de doi ani pentru „constanță”, „constanță a trăsăturilor”, pentru „puritate a sângelui” - în 1854 și în 1855. Experimentele cu mazăre au continuat timp de opt ani. De sute de ori în opt înflorind cu propriile mâini, a tăiat cu atenție anterele și, tastând polenul din staminele unei flori de altă varietate pe pensete, l-a aplicat pe stigmatul pistilului. Pentru zece mii de plante obținute ca urmare a încrucișărilor și de la hibrizi auto-polenizați, au fost introduse zece mii de pașapoarte. Mențiunile din ele sunt îngrijite: când planta mamă a fost cultivată, ce flori avea, al cărui polen a fost fertilizat, care mazăre - galben sau verde, neted sau ridat - au fost obținute, care flori - culoarea de la margini, culoarea în centrul - a înflorit când s-au primit semințele câte dintre ele sunt galbene, câte sunt verzi, rotunde, ridate, câte dintre ele sunt selectate pentru plantare, când sunt plantate și așa mai departe.
Rezultatul cercetărilor sale a fost raportul „Experimente pe hibrizi de plante”, care a fost citit de un naturalist brunnian în 1865. Raportul spune: „Motivul pentru înființarea experimentelor, căruia îi este dedicat acest articol, a fost încrucișarea artificială a plantelor ornamentale, care a fost efectuată pentru a obține forme noi, diferite prin culoare. Pentru înființarea unor experimente suplimentare pentru a urmări dezvoltarea hibrizilor în urmașii lor, au dat un impuls regularității evidente cu care formele hibride s-au întors constant la formele lor ancestrale. Așa cum se întâmplă adesea în istoria științei, opera lui Mendel nu a primit imediat recunoașterea cuvenită de la contemporanii săi. Rezultatele experimentelor sale au fost publicate la o reuniune a Societății de Științe Naturale din orașul Brunn și apoi publicate în jurnalul acestei societăți, dar ideile lui Mendel la acel moment nu au găsit sprijin. Un număr al revistei care descrie opera revoluționară a lui Mendel adună praf în biblioteci de treizeci de ani. Abia la sfârșitul secolului al XIX-lea, oamenii de știință care se ocupau de problemele eredității au descoperit operele lui Mendel și el a putut primi (deja postum) recunoașterea pe care o merita.
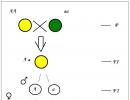
Prima lege a lui Mendel
Se numește traversarea a două organisme hibridizare, descendenții de la traversarea a doi indivizi cu ereditate diferită sunt numiți hibrid,și o persoană - un hibrid.Monohibrid se numește încrucișarea a două organisme care diferă unele de altele într-o pereche de trăsături alternative (care se exclud reciproc). În consecință, cu o astfel de încrucișare, sunt urmărite tiparele de moștenire a doar două trăsături, a căror dezvoltare se datorează unei perechi de gene alelice. Toate celelalte semne caracteristice acestor organisme nu sunt luate în considerare.
Dacă traversați plantele de mazăre cu semințe galbene și verzi, atunci toți hibrizii rezultați vor avea semințe galbene. Aceeași imagine se observă atunci când traversează plante cu o formă de semințe netedă și ridată; toți descendenții primei generații vor avea o formă netedă de semințe:
Prin urmare, a fost numită prima lege a lui Mendel Legea uniformității hibrizilor din prima generație.
Dacă folosim termenii care au apărut mulți ani după munca lui Mendel, atunci putem spune că celulele plantelor de mazăre dintr-un soi conțin două gene de numai culoare galbenă, iar genele plantelor dintr-o altă varietate - două gene de numai culoare verde. Genele responsabile de dezvoltarea unei trăsături (de exemplu, culoarea semințelor) sunt numite gene alelice... În consecință, într-un hibrid din prima generație, se dezvoltă doar una din fiecare pereche de trăsături alternative. Al doilea semn pare să dispară, nu apare. Fenomenul predominanței într-un hibrid cusemn al unuia dintre părinți G. Mendel numit dominraționarea. A fost numită o trăsătură care se manifestă într-un hibrid din prima generație și care suprimă dezvoltarea unei alte trăsături dominant,și opusul, adică suprimat, semn - recesiv. Gregor Mendel a decis să înlocuiască descrierea trăsăturilor plantelor cu codul abstract „A, B, C, D, E, F, G” și „a, b, c, d, e, f, g” și apoi de la observarea soarta unei perechi de trăsături a trecut la urmărirea a două, trei, patru perechi în același timp. Mare A, B, C, D, E, F, G, el a marcat trăsăturile dominante; mic a, b, c, d, e, f, g - recesiv. Dacă există două gene alelice identice în genotipul unui organism (zigot), ambele sunt dominante sau ambele sunt recesive (AA sau aa), un astfel de organism se numește homozigot. Dacă dintr-o pereche de gene alelice, una este dominantă, iar cealaltă este recesivă (Aa), atunci se numește un astfel de organism heterozigot.
Legea divizării sau a doua lege a lui Mendel.
Dacă descendenții primei generații, identici în trăsătura studiată, sunt încrucișați între ei, atunci în a doua generație trăsăturile ambilor părinți apar într-un anumit raport numeric: 3/4 dintre indivizi vor avea o trăsătură dominantă, ¼ recesiv:

Fenomenul în care traversarea heterozigoțilorindivizii duc la formarea descendenților, dintre care uniirogo poartă o trăsătură dominantă și parte- recesivny, se numește divizare.În consecință, trăsătura recesivă a hibrizilor din prima generație nu a dispărut, ci a fost doar suprimată și se va manifesta în a doua generație hibridă.
Ipoteza purității gametelor. Mendel a sugerat că în timpul formării hibrizilor, factorii ereditari nu se amestecă, dar rămân neschimbate. În hibrid, ambii factori sunt prezenți - dominanți și recesivi, dar factorul ereditar dominant se manifestă sub forma unei trăsături, în timp ce factorul recesiv este suprimat. Comunicarea între generații în timpul reproducerii sexuale se realizează prin intermediul celulelor sexuale - Hameta. Prin urmare, trebuie să presupunem că fiecare gamet are un singur factor dintr-o pereche. Apoi, în timpul fertilizării, fuziunea a doi gameți, fiecare dintre aceștia purtând un factor ereditar recesiv, va duce la formarea unui organism cu o trăsătură recesivă care se manifestă fenotipic. Fuziunea gametilor, fiecare dintre aceștia purtând un factor dominant sau doi gameti, dintre care unul conține un factor dominant și celălalt un factor recesiv, va duce la dezvoltarea unui organism cu o trăsătură dominantă.
Mendel a explicat împărțirea descendenților atunci când traversează indivizi heterozigoți prin faptul că gametii sunt genetic puri, adică poartă o singură genă dintr-o pereche alelică. Ipoteză(numit acum legea) puritategameti poate fi formulat după cum urmează: în timpul formării celulelor germinale, o singură genă dintr-o pereche alelică intră în fiecare gamet.
De ce și cum se întâmplă acest lucru? Se știe că fiecare celulă a corpului are exact același set diploid de cromozomi. Doi cromozomi omologi conțin două gene identice. Gametii "puri" genetic se formează după cum urmează: atunci când gametii masculi și feminini se îmbină, se obține un hibrid cu un set diploid (dublu) de cromozomi.
După cum se poate vedea din diagramă (Anexa 2), zigotul primește jumătate din cromozomi de la organismul patern și jumătate de la matern.
În procesul de formare a gameților într-un hibrid, cromozomii omologi în timpul primei diviziuni meiotice intră, de asemenea, în celule diferite.
Pentru această pereche alelică, se formează două varietăți de gameți. În timpul fertilizării, gametii care poartă aceleași sau diferite alele se întâlnesc accidental. Datorită probabilității statistice, cu un număr suficient de mare de gameți la descendenți, 25% din genotipuri vor fi homozigoți dominanți, 50% - heterozigoți, 25% - homozigoți recesivi, adică raportul 1AA: 2Aa: 1 aa.
În consecință, conform fenotipului, descendenții celei de-a doua generații în timpul încrucișării monohidri este distribuit într-un raport de 3: 1 (¾ indivizi cu trăsătură dominantă, ¼ indivizi cu trăsătură recesivă).
Baza citologică pentru clivarea caracterelor în încrucișarea monohibridă este divergența cromozomilor omologi la diferiți poli ai celulei și formarea celulelor germinale haploide în meioză.
În exemplele discutate mai sus, regula uniformității a fost exprimată prin faptul că toți hibrizii erau asemănători exterior cu unul dintre părinți. Nu este întotdeauna cazul. Adesea semnele din formele heterozigote sunt de natură intermediară, adică dominarea poate să nu fie completă. Schema de încrucișare a două forme ereditare ale plantei Night Beauty:


Una dintre ele are flori roșii (și aceasta este caracteristica dominantă), iar cealaltă este albă. Diagrama arată că toți hibrizii din prima generație au flori roz. În a doua generație, divizarea are loc într-un raport de 1: 2: 1, adică o floare roșie (homozigot), două flori roz (heterozigot), una albă (homozigot). Acest fenomen se numește dominanță incompletă.
În cazul unei dominații incomplete, gena dominantă într-o stare heterozigotă nu suprima întotdeauna complet gena recesivă. În unele cazuri, un hibrid fi nu reproduce pe deplin niciuna dintre trăsăturile părintești și trăsătura este de natură intermediară cu o abatere mai mult sau mai mică spre o stare dominantă sau recesivă. Dar toți indivizii acestei generații sunt uniformi în această caracteristică. Dominația incompletă este răspândită. S-a găsit în studiul moștenirii culorii florilor în snapdragons, a culorii lânii la bovine și ovine, trăsături biochimice la om etc. Trăsăturile intermediare care rezultă din dominarea incompletă reprezintă adesea valoare estetică sau materială pentru oameni. Se pune întrebarea: este posibil să se reproducă prin selecție, de exemplu, o varietate de frumusețe de noapte cu o culoare roz de flori? Evident că nu, deoarece această trăsătură se dezvoltă numai la heterozigoți și atunci când sunt încrucișați între ei, se produce întotdeauna divizarea:
Dominația incompletă este răspândită. S-a găsit în studiul moștenirii culorii florilor în snapdragons, a culorii lânii la bovine și ovine, trăsături biochimice la om etc. Trăsăturile intermediare care rezultă din dominarea incompletă reprezintă adesea valoare estetică sau materială pentru oameni. Se pune întrebarea: este posibil să se reproducă prin selecție, de exemplu, o varietate de frumusețe de noapte cu o culoare roz de flori? Evident că nu, deoarece această trăsătură se dezvoltă numai la heterozigoți și atunci când sunt încrucișați între ei, se produce întotdeauna divizarea.
Legea combinației independente sau a treiaLegea lui Mendel.
Studiul lui Mendel asupra moștenirii unei perechi de alele a făcut posibilă stabilirea unui număr de modele genetice importante: fenomenul dominanței, imuabilitatea alelelor recesive în hibrizi, împărțirea descendenței hibrizilor într-un raport de 3: 1, și, de asemenea, să presupunem că gametii sunt genetic puri, adică conțin o singură genă din perechi alelice. Cu toate acestea, organismele diferă în multe gene. Este posibil să se stabilească tiparele de moștenire a două perechi de trăsături alternative și mai mult de dihidrid sau traversare polibridă, adică încrucișarea formelor parentale diferind în două perechi de trăsături.
Pentru încrucișarea dihidridă, Mendel a luat plante de mazăre homozigote care diferă în doi indicatori - culoarea semințelor (galben, verde) și forma semințelor (netede, ridate). Caracteristici dominante - colorare galbenă (A)și formă netedă (V) semințe. Fiecare plantă formează o varietate de gameți pentru alelele studiate: Când gametele se unesc, toți descendenții vor fi uniformi:


|
Rețea de panetă |
||||||

Este ușor de calculat că, în funcție de fenotip, descendenții sunt împărțiți în 4 grupe: 9 galbene netede, 3 galbene ridate, 3 verzi netede, 1 galbene ridate (9: 3: 3: 1). Dacă luăm în considerare rezultatele împărțirii pentru fiecare pereche de caractere separat, se dovedește că raportul dintre numărul de semințe galbene și numărul de semințe verzi și raportul dintre semințele netede și semințele încrețite pentru fiecare pereche este de 3: 1 . Aceasta poate fi exprimată algebric ca pătratul binomului
(3 + 1) ² = 3² + 2 · 3 + 1² sau 9 + 3 + 3 + 1
Astfel, într-o încrucișare dihidridă, fiecare pereche de trăsături în timpul despărțirii în descendenți se comportă în același mod ca într-o încrucișare monohibridă, adică independent de cealaltă pereche de trăsături.
În timpul fertilizării, gametele sunt combinate conform regulilor combinațiilor aleatorii, dar cu probabilitate egală pentru fiecare. În zigotii rezultați apar diverse combinații de gene. Acum putem formula a treia lege a lui Mendel: la traversarea a doi indivizi homozigoti, diferițiseparate unele de altele de două sau mai multe perechi de trăsături alternative, gene șitrăsăturile lor corespunzătoare sunt moștenite independentunul de celălalt și sunt combinate în tot posibilulcombinații.
Legile lui Mendel servesc ca bază pentru analiza divizării în cazuri mai complexe: atunci când indivizii diferă în trei, patru perechi de caractere sau mai mult.
Condiții pentru respectarea legilor moștenirii Mendel
Legile descoperite de Gregor Mendel nu sunt întotdeauna aplicabile în genetică. Există multe condiții pentru respectarea legilor lui Mendel. Pentru astfel de cazuri, există alte legi (de exemplu: legea lui Morgan) sau explicații.
Să formulăm condițiile de bază pentru respectarea legilor moștenirii.
Pentru a respecta legea uniformității hibrizilor din prima generație, este necesar ca:
organismele parentale erau homozigote;
genele diferitelor alele au fost localizate în cromozomi diferiți și nu într-unul singur (în caz contrar, poate apărea fenomenul „moștenirii legate”).
Legea divizării va fi respectată dacă
la hibrizi, factorii ereditari rămân neschimbați;
Legea distribuției independente a genelor la descendenți și apariția diferitelor combinații ale acestor gene în timpul încrucișării dihidri este posibilă doar în caz.
dacă perechile de gene alelice sunt situate în perechi diferite de cromozomi omologi.
Încălcarea acestor condiții poate duce fie la absența divizării în a doua generație, fie la divizarea în prima generație; sau la denaturarea raportului dintre diferite genotipuri și fenotipuri. Legile lui Mendel sunt universale pentru toate organismele diploide care se reproduc sexual. În general, acestea sunt valabile pentru gene autozomale cu penetranță completă (frecvență de manifestare 100% a trăsăturii analizate; penetranță 100% implică faptul că trăsătura este exprimată în toți purtătorii alelei care determină dezvoltarea acestei trăsături) și expresivitate constantă; expresivitatea constantă implică faptul că severitatea fenotipică a unei trăsături este aceeași sau aproximativ aceeași la toți purtătorii alelei care determină dezvoltarea acestei trăsături.
Concluzie.
Legile lui Gregor Mendel sunt utilizate în prezent pe scară largă în reproducerea plantelor, animalelor și microorganismelor, în medicină, inginerie genetică și multe alte ramuri ale vieții umane, precum și în rezolvarea problemelor din genetică.
Este important de reținut că Mendel a formulat legi și a tras concluzii într-un moment în care nu se știa nimic despre ADN, gene și cromozomi. Cu toate acestea, s-a dovedit a fi complet drept și, deși nu imediat, teoriile sale au fost recunoscute și luate ca bază a științei în dezvoltare a geneticii.
Teoria mendeliană a eredității, adică totalitatea ideilor despre determinanții ereditari și natura transmiterii lor de la părinți la descendenți, în sensul său, este direct opusă teoriilor domenialeene, în special, teoria pangenezei propusă de Darwin. În conformitate cu această teorie, semnele părinților sunt directe, adică din toate părțile corpului, transmise descendenților. Prin urmare, caracterul trăsăturii copilului ar trebui să depindă direct de proprietățile părintelui. Acest lucru contrazice complet concluziile trase de Mendel: factorii determinanți ai eredității, adică genele sunt prezente în organism relativ independent de sine. Natura trăsăturilor (fenotipul) este determinată de combinația lor aleatorie. Ele nu sunt modificate de nicio parte a corpului și se află într-o relație dominanță-recesivă. Astfel, teoria mendeliană a eredității se opune ideii de moștenire a caracterelor dobândite în timpul dezvoltării individuale.
Experimentele lui Mendel au servit ca bază pentru dezvoltarea geneticii moderne - o știință care studiază două proprietăți principale ale unui organism - ereditatea și variabilitatea. El a reușit să identifice tiparele de moștenire grație unor abordări metodologice fundamental noi:
1) Mendel a ales cu succes obiectul cercetării;
2) a analizat moștenirea trăsăturilor individuale în urmașii plantelor încrucișate care diferă în una, două și trei perechi de trăsături alternative contrastante. În fiecare generație, o evidență a fost ținută separat pentru fiecare pereche de aceste caracteristici;
3) el a înregistrat nu numai rezultatele obținute, ci și a efectuat prelucrarea lor matematică.
Metodele simple de cercetare enumerate au constituit o metodă hibridologică fundamental nouă pentru studierea moștenirii, care a devenit baza pentru cercetări suplimentare în genetică.
Bibliografie.
Biologie generală: Manual pentru clasele 9-10. Miercuri școală / Polyansky Yu.I., Brown A.D., Verzilin N.M. și alții; M.: Educație, 1987.-287 p.: Bolnav.
Mendel a ajuns la concluzia că predispoziția la naștere ... afectează frecvențele genotipurilor din populație. Legile Mendel nu ne spune nimic despre frecvențe ...Prima lege a lui Mendel
Se numește traversarea a două organisme hibridizare, descendenții de la traversarea a doi indivizi cu ereditate diferită sunt numiți hibrid,și o persoană - un hibrid. Mono hibrid se numește încrucișarea a două organisme care diferă unele de altele într-o pereche de trăsături alternative (care se exclud reciproc). În consecință, cu o astfel de încrucișare, sunt urmărite tiparele de moștenire a doar două trăsături, a căror dezvoltare se datorează unei perechi de gene alelice. Toate celelalte semne caracteristice acestor organisme nu sunt luate în considerare.
Dacă traversați plantele de mazăre cu semințe galbene și verzi, atunci toți hibrizii rezultați vor avea semințe galbene. Aceeași imagine se observă atunci când traversează plante cu o formă de semințe netedă și ridată; toți descendenții primei generații vor avea o formă netedă de semințe:
Prin urmare, a fost numită prima lege a lui Mendel Legea uniformității hibrizilor din prima generație.
Dacă folosim termenii care au apărut mulți ani după munca lui Mendel, atunci putem spune că celulele plantelor de mazăre dintr-un soi conțin două gene de numai culoare galbenă, iar genele plantelor dintr-o altă varietate - două gene de numai culoare verde. Genele responsabile de dezvoltarea unei trăsături (de exemplu, culoarea semințelor) sunt numite gene alelice... În consecință, într-un hibrid din prima generație, se dezvoltă doar una din fiecare pereche de trăsături alternative. Al doilea semn pare să dispară, nu apare. Fenomenul predominanței într-un hibrid cu semn al unuia dintre părinți G. Mendel numit domin raționarea. A fost numită o trăsătură care se manifestă într-un hibrid din prima generație și care suprimă dezvoltarea unei alte trăsături dominant,și opusul, adică suprimat, semn - recesiv. Gregor Mendel a decis să înlocuiască descrierea trăsăturilor plantelor cu codul abstract „A, B, C, D, E, F, G” și „a, b, c, d, e, f, g” și apoi de la observarea soarta unei perechi de trăsături a trecut la urmărirea a două, trei, patru perechi în același timp. Mare A, B, C, D, E, F, G, el a marcat trăsăturile dominante; mic a, b, c, d, e, f, g - recesiv. Dacă există două gene alelice identice în genotipul unui organism (zigot), ambele sunt dominante sau ambele sunt recesive (AA sau aa), un astfel de organism se numește homozigot. Dacă dintr-o pereche de gene alelice, una este dominantă, iar cealaltă este recesivă (Aa), atunci se numește un astfel de organism heterozigot.
Legea divizării sau a doua lege a lui Mendel.
Dacă descendenții primei generații, identici în trăsătura studiată, sunt încrucișați între ei, atunci în a doua generație trăsăturile ambilor părinți apar într-un anumit raport numeric: 3/4 dintre indivizi vor avea o trăsătură dominantă, ¼ recesiv:

Fenomenul în care traversarea heterozigoților indivizii duc la formarea descendenților, dintre care unii rogo poartă o trăsătură dominantă și parte - recesiv ny, se numește divizare.În consecință, trăsătura recesivă a hibrizilor din prima generație nu a dispărut, ci a fost doar suprimată și se va manifesta în a doua generație hibridă.
Ipoteza purității gametelor. Mendel a sugerat că în timpul formării hibrizilor, factorii ereditari nu se amestecă, dar rămân neschimbate. În hibrid, ambii factori sunt prezenți - dominanți și recesivi, dar factorul ereditar dominant se manifestă sub forma unei trăsături, în timp ce factorul recesiv este suprimat. Comunicarea între generații în timpul reproducerii sexuale se realizează prin intermediul celulelor sexuale - Ha meta. Prin urmare, trebuie să presupunem că fiecare gamet are un singur factor dintr-o pereche. Apoi, în timpul fertilizării, fuziunea a doi gameți, fiecare dintre aceștia purtând un factor ereditar recesiv, va duce la formarea unui organism cu o trăsătură recesivă care se manifestă fenotipic. Fuziunea gametilor, fiecare dintre aceștia purtând un factor dominant sau doi gameti, dintre care unul conține un factor dominant și celălalt un factor recesiv, va duce la dezvoltarea unui organism cu o trăsătură dominantă.
Mendel a explicat împărțirea descendenților atunci când traversează indivizi heterozigoți prin faptul că gametii sunt genetic puri, adică poartă o singură genă dintr-o pereche alelică. Ipoteză(numit acum legea) puritate gameti poate fi formulat după cum urmează: în timpul formării celulelor germinale, o singură genă dintr-o pereche alelică intră în fiecare gamet.
De ce și cum se întâmplă acest lucru? Se știe că fiecare celulă a corpului are exact același set diploid de cromozomi. Doi cromozomi omologi conțin două gene identice. Gametii "puri" genetic se formează după cum urmează: atunci când gametii masculi și feminini se îmbină, se obține un hibrid cu un set diploid (dublu) de cromozomi.
După cum se poate vedea din diagramă (Anexa 2), zigotul primește jumătate din cromozomi de la organismul patern și jumătate de la matern.
În procesul de formare a gameților într-un hibrid, cromozomii omologi în timpul primei diviziuni meiotice intră, de asemenea, în celule diferite.
Pentru această pereche alelică, se formează două varietăți de gameți. În timpul fertilizării, gametii care poartă aceleași sau diferite alele se întâlnesc accidental. Datorită probabilității statistice, cu un număr suficient de mare de gameți la descendenți, 25% din genotipuri vor fi homozigoți dominanți, 50% - heterozigoți, 25% - homozigoți recesivi, adică raportul 1AA: 2Aa: 1 aa.
În consecință, conform fenotipului, descendenții celei de-a doua generații în timpul încrucișării monohidri este distribuit într-un raport de 3: 1 (¾ indivizi cu trăsătură dominantă, ¼ indivizi cu trăsătură recesivă).
Baza citologică pentru clivarea caracterelor în încrucișarea monohibridă este divergența cromozomilor omologi la diferiți poli ai celulei și formarea celulelor germinale haploide în meioză.
În exemplele discutate mai sus, regula uniformității a fost exprimată prin faptul că toți hibrizii erau asemănători exterior cu unul dintre părinți. Nu este întotdeauna cazul. Adesea semnele din formele heterozigote sunt de natură intermediară, adică dominarea poate să nu fie completă. Schema de încrucișare a două forme ereditare ale plantei Night Beauty:


Una dintre ele are flori roșii (și aceasta este caracteristica dominantă), iar cealaltă este albă. Diagrama arată că toți hibrizii din prima generație au flori roz. În a doua generație, divizarea are loc într-un raport de 1: 2: 1, adică o floare roșie (homozigot), două flori roz (heterozigot), una albă (homozigot). Acest fenomen se numește dominanță incompletă.
În cazul unei dominații incomplete, gena dominantă într-o stare heterozigotă nu suprima întotdeauna complet gena recesivă. În unele cazuri, un hibrid fi nu reproduce pe deplin niciuna dintre trăsăturile părintești și trăsătura este de natură intermediară cu o abatere mai mult sau mai mică spre o stare dominantă sau recesivă. Dar toți indivizii acestei generații sunt uniformi în această caracteristică. Dominația incompletă este răspândită. S-a găsit în studiul moștenirii culorii florilor în snapdragons, a culorii lânii la bovine și ovine, trăsături biochimice la om etc. Trăsăturile intermediare care rezultă din dominarea incompletă reprezintă adesea valoare estetică sau materială pentru oameni. Se pune întrebarea: este posibil să se reproducă prin selecție, de exemplu, o varietate de frumusețe de noapte cu o culoare roz de flori? Evident că nu, deoarece această trăsătură se dezvoltă numai la heterozigoți și atunci când sunt încrucișați între ei, se produce întotdeauna divizarea:
Dominația incompletă este răspândită. S-a găsit în studiul moștenirii culorii florilor în snapdragons, a culorii lânii la bovine și ovine, trăsături biochimice la om etc. Trăsăturile intermediare care rezultă din dominarea incompletă reprezintă adesea valoare estetică sau materială pentru oameni. Se pune întrebarea: este posibil să se reproducă prin selecție, de exemplu, o varietate de frumusețe de noapte cu o culoare roz de flori? Evident că nu, deoarece această trăsătură se dezvoltă numai la heterozigoți și atunci când sunt încrucișați între ei, se produce întotdeauna divizarea.
Legea combinației independente sau a treia Legea lui Mendel.
Studiul lui Mendel despre moștenirea unei perechi de alele a făcut posibilă stabilirea unui număr de modele genetice importante: fenomenul dominanței, imuabilitatea alelelor recesive în hibrizi, împărțirea descendenței hibrizilor într-un raport de 3: 1, și, de asemenea, să presupunem că gametii sunt genetic puri, adică conțin o singură genă din perechi alelice. Cu toate acestea, organismele diferă în multe gene. Este posibil să se stabilească tiparele de moștenire a două perechi de trăsături alternative și mai mult de dihidrid sau traversare polibridă, adică încrucișarea formelor parentale diferind în două perechi de trăsături.
Un om de știință din Republica Cehă Gregor Mendel (1822-1884), în cursul cercetărilor sale privind încrucișarea diferitelor soiuri de mazăre, a dezvăluit anumite tipare care sunt acum cunoscute pe scară largă sub numele celor trei legi ale lui Mendel.
Prima lege
Dacă exemplarele de plante au fost obținute în prima generație prin încrucișarea soiurilor parentale homozigote (linii pure) cu trăsături diferite, atunci descendenții sunt aceiași în genotip și fenotip. În acest caz, o trăsătură este înțeleasă ca orice calitate selectată a plantelor, prin care se pot distinge două soiuri. Prima lege a lui Mendel este cunoscută și ca regula dominanței sau legea uniformității pentru hibrizii din prima generație. Trăsătura identificată în prima generație a fost numită dominantă, iar calitatea care a fost suprimată și care nu s-a manifestat a fost recesivă.
A doua lege
Legea divizării. Esența sa constă în faptul că la traversarea, de exemplu, în timpul autopolenizării, a aceluiași tip de hibrizi din prima generație din generația următoare, descendenții sunt împărțiți în funcție de trăsătura aleasă (fenotip) într-un raport de 3 : 1 în cazul dominanței complete și 1: 2: 1 în cazul dominanței incomplete.
A treia lege
A treia lege a lui Mendel se referă la moștenirea independentă a trăsăturilor sau la combinația independentă a genelor plantelor. Dacă există o încrucișare de exemplare de plante care diferă între ele în două sau mai multe perechi de trăsături alternative, materialul genetic și trăsăturile fenotipice pe care le determină sunt moștenite independent unul de celălalt și pot fi combinate în orice combinație.
Experimentele omului de știință ceh au devenit baza dezvoltării geneticii moderne. G. Mendel a reușit să descopere tiparele de bază ale moștenirii cu ajutorul celor mai recente abordări metodologice, care sunt încă folosite de toți geneticienii.
În experimentele sale, toate exemplarele încrucișate aparțineau aceleiași specii de plante. Este important ca, în același timp, să se deosebească în mod clar unul de celălalt într-una, două, mai multe perechi de semne alternative (opuse). Aceste trăsături au fost constante, adică au fost transmise de la generație la generație atunci când traversau în aceeași specie. A fost necesar să se studieze descendenții din fiecare pereche de hibrizi în mod individual și să se facă o prezentare cantitativă a hibrizilor care diferă în anumite perechi de trăsături alternative.
Aceste metode de cercetare au stat la baza unei noi metode hibridologice, cu apariția căreia a început epoca studiului eredității și variabilității.
Modelele de moștenire a trăsăturilor în timpul reproducerii sexuale au fost stabilite de G. Mendel. Este necesar să se înțeleagă clar genotipul și fenotipul, alelele, homo- și heterozigoza, dominanța și tipurile sale, tipurile de încrucișări și, de asemenea, să întocmească scheme.
Monohibrid se numește încrucișare, în care formele părintești diferă între ele într-o pereche de caractere alternative, contrastante.
În consecință, cu o astfel de încrucișare, sunt urmărite tiparele de moștenire a doar două variante ale trăsăturii, a căror dezvoltare se datorează unei perechi de gene alelice. Exemple de încrucișări monohidri efectuate de G. Mendel sunt cruci de mazăre cu caractere alternative clar vizibile precum florile violete și albe, culoarea galbenă și verde a fructelor imature (fasole), suprafața netedă și încrețită a semințelor, culoarea galbenă și verde etc.
Uniformitatea hibrizilor din prima generație (prima lege a lui Mendel) Atunci când traversează mazărea cu flori violete (AA) și albe (aa), Mendel a constatat că toate plantele hibride din prima generație (F1) aveau flori violete (Figura 2).
Imaginea 2 – Schema de trecere monohibridă
În același timp, culoarea albă a florii nu a apărut. Atunci când traversează plante care au semințe netede și ridate, hibrizii vor avea semințe netede. G. Mendel a constatat, de asemenea, că toți hibrizii F 1 erau uniformi (omogeni) pentru fiecare dintre cele șapte trăsături pe care le-a studiat. În consecință, la hibrizii din prima generație, dintr-o pereche de trăsături alternative părintești, apare doar una, iar trăsătura celuilalt părinte pare să dispară.
Semnele alternative sunt semne care se exclud reciproc, contrastante.
Mendel a numit predominanța trăsăturilor unuia dintre părinți în hibrizii F1 ca dominanță, iar trăsătura corespunzătoare ca dominantă. El a numit trăsăturile care nu apar în hibrizii F 1 recesivi. Deoarece toți hibrizii din prima generație sunt uniformi, acest fenomen a fost numit primele legi ale lui Mendel, sau legea uniformității hibride din prima generație și regula dominanței.
Poate fi formulat după cum urmează: atunci când traversează două organisme aparținând unor linii pure diferite (două organisme homozigote), care diferă între ele într-o pereche de trăsături alternative, întreaga primă generație de hibrizi va fi uniformă și va purta trăsătura uneia dintre parintii.
Fiecare genă are două stări - „A” și „a”, deci alcătuiesc o pereche și fiecare membru al perechii se numește alelă. Genele situate în aceleași loci (secțiuni) ale cromozomilor omologi și care determină dezvoltarea alternativă a aceleiași trăsături se numesc alelice.
De exemplu, culoarea purpurie și albă a unei flori de mazăre este dominantă și respectiv recesivă pentru două alele (A și a) ale unei gene. Datorită prezenței a două alele, sunt posibile două stări ale organismului: homo- și heterozigoți. Dacă un organism conține aceleași alele ale unei anumite gene (AA sau aa), atunci se numește homozigot pentru această genă (sau trăsătură), iar dacă este diferit (Aa), atunci heterozigot. Prin urmare, o alelă este o formă a existenței unei gene. Un exemplu de genă tri-alelică este o genă care determină sistemul de grupare sanguină ABO la om. Există mai multe alele: pentru gena care controlează sinteza hemoglobinei umane, există zeci dintre ele.
Din semințele de mazăre hibride, Mendel a crescut plante, pe care le-a supus autopolenizării, iar semințele rezultate au fost semănate din nou. Rezultatul a fost o a doua generație de hibrizi, sau hibrizi F 2. Dintre acestea din urmă, s-a găsit o despărțire pentru fiecare pereche de trăsături alternative într-un raport de aproximativ 3: 1, adică trei sferturi din plante aveau trăsături dominante (flori purpurii, semințe galbene, semințe netede etc.) și un sfert erau recesiv (flori albe, semințe verzi, semințe ridate etc.). În consecință, trăsătura recesivă din hibridul F1 nu a dispărut, ci a fost suprimată și reapărută doar în a doua generație. Această generalizare a fost numită ulterior a doua lege a lui Mendel sau legea divizării.
Scindarea este un fenomen în care încrucișarea indivizilor heterozigoți duce la formarea descendenților, dintre care unii sunt dominanți, iar alții sunt recesivi.
A doua lege a lui Mendel: atunci când doi descendenți din prima generație sunt încrucișați între ei (doi indivizi heterozigoți) în a doua generație, divizarea se observă într-un anumit raport numeric: conform fenotipului 3: 1, conform genotipului 1: 2 : 1 (Fig. 3).

Figura 3 - Schema semnelor de divizare
la traversarea hibrizilor F 1
Împărțirea trăsăturilor la descendenți atunci când traversează indivizi heterozigoți a fost explicată de G. Mendel prin faptul că gametii sunt genetic puri, adică poartă o singură genă dintr-o pereche alelică. Legea purității gametelor poate fi formulată după cum urmează: în timpul formării celulelor germinale, o singură genă dintr-o pereche alelică intră în fiecare gamet.
Trebuie avut în vedere faptul că utilizarea metodei hibridologice pentru a analiza moștenirea trăsăturilor la orice specie de animale sau plante implică următoarele încrucișări:
încrucișarea formelor parentale (P), diferind într-o singură (încrucișare monohibridă) sau mai multe perechi (încrucișare polibridă) de trăsături alternative și obținerea hibrizilor din prima generație (F 1);
traversarea hibrizilor F 1 între ei și obținerea hibrizilor din a doua generație (F 2);
analiza matematică a rezultatelor traversării.
Ulterior, Mendel s-a orientat spre studiul trecerii cu dihidri.
Traversare dihidridă Este o încrucișare în care sunt implicate două perechi de alele (genele perechi sunt alelice și sunt localizate doar în cromozomii omologi).
În timpul încrucișării dihidri, G. Mendel a studiat moștenirea trăsăturilor pentru care sunt responsabile genele care se află în diferite perechi de cromozomi omologi. În acest sens, fiecare gamet trebuie să conțină o genă din fiecare pereche alelică.
Hibrizii heterozigoți pentru două gene se numesc diheterozigoți și, dacă diferă în trei sau mai multe gene, sunt numiți tri- și, respectiv, poliheterozigoți.
Scheme mai complexe de încrucișări dihidri, înregistrarea genotipurilor și fenotipurilor F2 se efectuează folosind rețeaua Pennett. Luați în considerare un exemplu de astfel de cruce. Pentru încrucișare, au fost luate două forme parentale inițiale homozigote: prima formă avea semințe galbene și netede; a doua formă avea semințe verzi și ridate (Fig. 4).

Figura 4 - Traversarea dihidridă a plantelor de mazăre,
diferite ca formă și culoare semințelor
Culoarea galbenă și semințele netede sunt trăsături dominante; culoarea verde și semințele ridate sunt simptome recesive. Hibrizi de prima generație - încrucișați între ei. În a doua generație, scindarea fenotipică a fost observată într-un raport de 9: 3: 3: 1 sau (3 + 1) 2, după autopolenizarea hibrizilor F1, în conformitate cu legea scindării, semințele ridate și verzi au reapărut.
Plantele părinte în acest caz au genotipurile AABB și aabb, iar genotipul hibrizilor F 1 - AaBb, adică, este diheterozigot.
Astfel, atunci când traversează indivizi heterozigoți diferiți în mai multe perechi de trăsături alternative, scindarea fenotipului se observă la descendenți în raportul (3 + 1) n, unde n este numărul de perechi de trăsături alternative.
Genele care determină dezvoltarea diferitelor perechi de trăsături sunt numite non-alelice.
Rezultatele încrucișării dihidri și polihibri depind de dacă genele care determină trăsăturile luate în considerare sunt localizate în același cromozom sau în cromozomi diferiți. Mendel a dat peste trăsături ale căror gene se găseau în perechi diferite de cromozomi omologi de mazăre.
În meioză, cromozomii omologi de diferite perechi sunt combinați aleatoriu în gameți. Dacă cromozomul patern al primei perechi intră în gamet, atunci cu o probabilitate egală, atât cromozomul patern, cât și cel matern al celei de-a doua perechi pot intra în acest gamet. Prin urmare, trăsăturile ale căror gene se află în perechi diferite de cromozomi omologi sunt combinate independent una de cealaltă. Ulterior, sa dovedit că din cele șapte perechi de trăsături studiate de Mendel în mazăre, în care numărul diploid de cromozomi este de 2 n = 14, genele responsabile pentru una dintre perechile de trăsături se aflau pe același cromozom. Cu toate acestea, Mendel nu a găsit o încălcare a legii moștenirii independente, deoarece legătura dintre aceste gene nu a fost observată din cauza distanței mari dintre ele).
Pe baza cercetărilor sale, Mendel a dedus o a treia lege - legea moștenirii independente a trăsăturilor sau combinația independentă de gene.
Fiecare pereche de gene alelice (și trăsături alternative controlate de acestea) este moștenită independent una de cealaltă.
Legea combinării independente a genelor formează baza variabilității combinative observate în timpul încrucișării în toate organismele vii. De asemenea, observăm că, spre deosebire de prima lege a lui Mendel, care este întotdeauna adevărată, a doua lege este valabilă doar pentru gene localizate în diferite perechi de cromozomi omologi. Acest lucru se datorează faptului că cromozomii neomologi sunt combinați în celulă independent unul de celălalt, lucru dovedit nu numai prin studierea naturii moștenirii caracterelor, ci și printr-o metodă citologică directă.
Când studiați materialul, acordați atenție cazurilor de încălcări ale divizării regulate de către fenotip cauzate de acțiunea letală a genelor individuale.
Ereditate și variabilitate. Ereditatea și variabilitatea sunt cele mai importante proprietăți caracteristice tuturor organismelor vii.
Variabilitatea ereditară sau genotipică este subdivizată în combinativă și mutațională.
Variabilitatea se numește combinațională, care se bazează pe formarea recombinărilor, adică astfel de combinații de gene pe care părinții nu le-au avut.
Variabilitatea combinativă se bazează pe reproducerea sexuală a organismelor, în urma căreia apare o mare varietate de genotipuri. Trei procese sunt practic surse nelimitate de variație genetică:
Divergența independentă a cromozomilor omologi în prima diviziune meiotică. Combinația independentă de cromozomi din meioză este baza celei de-a treia legi a lui G. Mendel. Apariția semințelor de mazăre ridate, verzi, netede și galbene, în a doua generație, de la plantele încrucișate cu semințe ridate netede, galbene și verzi, este un exemplu de variație combinativă.
Schimb reciproc de regiuni ale cromozomilor omologi, sau încrucișare. Creează noi grupuri de legături, adică servește ca o sursă importantă de recombinare genetică a alelelor. Odată ajuns în zigot, cromozomii recombinați contribuie la apariția unor trăsături atipice pentru fiecare dintre părinți.
Combinație aleatorie de gameți în timpul fertilizării.
Aceste surse de variabilitate combinativă acționează independent și simultan, oferind în același timp o „amestecare” constantă a genelor, ceea ce duce la apariția organismelor cu un genotip și fenotip diferit (genele în sine nu se schimbă). Cu toate acestea, noi combinații de gene se descompun destul de ușor atunci când sunt transmise din generație în generație.
Un exemplu de variabilitate combinativă. Floarea de frumusețe de noapte are o genă pentru culoarea roșie a petalelor A și o genă pentru albul a. Corpul lui Aa are petale roz. Astfel, frumusețea nopții nu are o genă roz; rozul apare atunci când o combinație (combinație) a unei gene roșii și albe.
Persoana are o boală ereditară anemie cu celule secera. AA este norma, aa este moartea, Aa este SKA. Cu SKA, o persoană nu poate tolera un efort fizic crescut, în timp ce nu suferă de malarie, adică agentul cauzator al malariei Malaria Plasmodium nu se poate hrăni cu hemoglobina greșită. Această caracteristică este utilă în centura ecuatorială; nu există nicio genă pentru aceasta, aceasta apare dintr-o combinație de gene A și a.
Astfel, variația ereditară este sporită de variația combinativă. Apărute, mutațiile individuale se găsesc în vecinătatea altor mutații, fac parte din noi genotipuri, adică apar multe combinații de alele. Orice individ este unic genetic (cu excepția gemenilor identici și a indivizilor care au apărut datorită reproducerii asexuale a unei clone cu o singură celulă ca strămoș). Deci, dacă presupunem că în fiecare pereche de cromozomi omologi există o singură pereche de gene alelice, atunci pentru o persoană al cărei set haploid de cromozomi este 23, numărul de genotipuri posibile va fi de 3 până la 23 de grade. Un număr atât de mare de genotipuri este de 20 de ori mai mare decât toți oamenii de pe Pământ. Cu toate acestea, în realitate, cromozomii omologi diferă în mai multe gene, iar calculul nu ia în considerare fenomenul de încrucișare. . Prin urmare, numărul de genotipuri posibile este exprimat în numere astronomice și se poate afirma cu încredere că apariția a două persoane identice este practic improbabilă (cu excepția gemenilor identici, care au apărut din același ou fertilizat). Prin urmare, în special, urmează posibilitatea unei identificări fiabile a persoanei de către resturile de țesuturi vii, confirmarea sau excluderea paternității.
Astfel, schimbul de gene datorat încrucișării cromozomilor în prima diviziune a meiozei, recombinarea independentă și aleatorie a cromozomilor în meioză și șansa de fuziune a gametilor în procesul sexual sunt trei factori care asigură existența variabilității combinative. Variabilitatea mutațională a genotipului în sine.
Mutațiile sunt modificări bruște, moștenite ale materialului genetic, ducând la o schimbare a anumitor caracteristici ale unui organism.
Principalele prevederi ale teoriei mutaționale au fost elaborate de omul de știință G. De Vries în 1901 – 1903 și rezumați la aceasta:
Mutațiile apar brusc, în salturi, ca modificări discrete ale trăsăturilor;
Spre deosebire de modificările non-ereditare, mutațiile sunt schimbări calitative care se transmit din generație în generație;
Mutațiile se manifestă în moduri diferite și pot fi atât benefice, cât și dăunătoare, atât dominante, cât și recesive;
Probabilitatea de a detecta mutații depinde de numărul de indivizi examinați;
Mutații similare pot reapărea;
Mutațiile sunt nedirecționale (spontane), adică orice parte a cromozomului poate muta, provocând modificări atât în semnele minore, cât și în cele vitale.
Aproape orice modificare a structurii sau a numărului de cromozomi, în care celula își păstrează capacitatea de a se reproduce, provoacă o modificare ereditară a caracteristicilor organismului.
Prin natura schimbării genomului, adică setul de gene conținute în setul haploid de cromozomi, se disting mutațiile genice, cromozomiale și genomice.
Mutațiile genice sau punctuale sunt rezultatul unei modificări a secvenței de nucleotide dintr-o moleculă de ADN din cadrul unei gene.
O astfel de modificare a genei este reprodusă în timpul transcrierii în structura i-ARN; duce la o modificare a secvenței aminoacizilor din lanțul polipeptidic format în timpul translației pe ribozomi. Ca urmare, se sintetizează o altă proteină, ceea ce duce la o modificare a caracteristicii corespunzătoare a organismului. Acesta este cel mai frecvent tip de mutație și cea mai importantă sursă de variație ereditară a organismelor.
Mutațiile cromozomiale (rearanjări sau aberații) sunt modificări ale structurii cromozomilor care pot fi identificate și studiate la microscopul cu lumină.
Se cunosc rearanjamente de diferite tipuri:
o lipsa de – pierderea capetelor cromozomului;
Ștergere – pierderea unei porțiuni a cromozomului în partea sa mijlocie;
Duplicare – repetarea două sau multiple a genelor localizate într-o anumită parte a cromozomului;
Inversia – rotația secțiunii cromozomiale cu 180 °, în urma căreia genele din această secțiune sunt situate în secvența opusă celei obișnuite;
Translocare – schimbarea poziției oricărei părți a cromozomului în setul de cromozomi. Cel mai comun tip de translocație este reciproc, în care zonele sunt schimbate între doi cromozomi neomologi. O secțiune a unui cromozom își poate schimba poziția fără schimb reciproc, rămânând în același cromozom sau fiind inclusă în altele.
Mutații genomice - o modificare a numărului de cromozomi din genomul celulelor dintr-un organism. Acest fenomen apare în două direcții: către o creștere a numărului de seturi haploide întregi (poliploidie) și către pierderea sau includerea cromozomilor individuali (aneuploidie).
Poliploidie – o creștere multiplă a setului haploid de cromozomi. Celulele cu un număr diferit de seturi haploide de cromozomi sunt numite triploid (3 n), tetraploid (4 n), hexaploid (6 n), octaploid (8 n), etc. polii celulari în timpul meiozei sunt perturbați sau mitoză. Poliploidia duce la o schimbare a caracteristicilor unui organism și, prin urmare, este o sursă importantă de variabilitate în evoluție și selecție, în special la plante. Acest lucru se datorează faptului că hermafroditismul (auto-polenizarea), apomixis (partenogeneză) și reproducerea vegetativă sunt foarte răspândite în organismele vegetale. Prin urmare, aproximativ o treime din speciile de plante comune pe planeta noastră, – poliploide și, în condițiile puternic continentale ale Pamirului de munte înalt, cresc până la 85% din poliploide. Aproape toate plantele cultivate sunt, de asemenea, poliploide, care, spre deosebire de rudele lor sălbatice, au flori, fructe și semințe mai mari, iar mai mulți nutrienți se acumulează în organele de depozitare (tulpină, tuberculi). Poliploizii se adaptează mai ușor la condiții de viață nefavorabile, tolerează temperaturile scăzute și seceta mai ușor. De aceea sunt răspândite în regiunile de munte nordice și înalte.