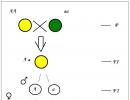
Schéma získavania hybridov prvej generácie. Mendelov prvý zákon je zákon nadvlády. Zákon čistoty gamét a jeho cytologické zdôvodnenie
6. Čo je ontogenéza. Aké sú obdobia ontogenézy? Stručne popíšte postembryonický vývoj.
1. Úrovne organizácií pre voľne žijúce zvieratá (zoznam).
2. Pomenujte organely dvoch membránových a nemembránových buniek. Uveďte vlastnosti ich štruktúry a funkcie.
3. Aké sú výhody procesu sexuálnej reprodukcie oproti asexuálu.
4. Tretí Mendelov zákon. Uveďte pomer potomstva podľa genotypu a fenotypu.
5. Vymenujte strediská pôvodu a rozmanitosti pestovaných rastlín podľa Vavilova. Kde je uložená zbierka semien rastlín rastúcich na Zemi, aký je jej význam.
6. Čo je ontogenéza. Aké sú obdobia ontogenézy? Stručne popíšte postembryonický vývoj.
telo hostiteľa. Preventívne opatrenia.
2. Hnacie sily evolúcie a ich vzťah.
5. Určte zväčšenie mikroskopu. Pripravte ho na prácu.
Lístok číslo 6
1. Obehový systém stavovcov.
2.1. Monohybridný prechod.
2. Prvý a druhý Mendelov zákon.
6. Použitím herbárového materiálu zoskupte rastliny do rodín.
Lístok číslo 7
1. Oddelenia nervového systému: centrálny a periférny. 2. Biotop organizmov.
Biológia, pomôžte !!!1. Veda, ktorá študuje vzorce prenosu znakov dedičnosťou
2. Najprv sa sformulovali genetické vzorce
3. Spárované chromozómy, z ktorých každý ide do tela od jedného z rodičov
4. Chromozómy obsahujúce súbor identických génov
5. Chromozómy umiestnené v tom istom organizme, ale líšia sa veľkosťou, tvarom a sadou génov
6. Gény umiestnené v rovnakých oblastiach homológnych chromozómov, zodpovedné za syntézu rovnakých proteínov, ale s odlišnou nukleotidovou sekvenciou
7. Aký bude názov génov kódujúcich ten istý proteín (napríklad hemoglobín alebo melanín), ktoré však budú obsahovať inú aminokyselinovú sekvenciu
8. Pri monohybridnom krížení sa značky skúmajú
9. Podľa génu pre farby (v Mendelových experimentoch) mal hrach počet znakov
10. Zygota, ktorá dostala dve rôzne alely
11. Zygote, ktorá dostala obe rovnaké alely
12. Gén označený krížením malým písmenom
13. Ako sa volá alela, označená pri krížení veľkým (veľkým) písmenom
14. Alela, ktorá sa fenotypicky objavuje iba v homozygotnom stave
15. Alela, ktorá sa navonok prejavuje v každom prípade, ak je vôbec prítomná v genotype jedinca
16. Zygote genotyp označený AA
17. Genotyp zygoty označený Aa
18. Genotyp zygoty, označený ako aa
19. Súbor vonkajších znakov tela
20. Súhrn všetkých dedičných charakteristík organizmu
21. Súbor všetkých génov druhu alebo populácie
22. Rozdelenie hybridov v druhej generácii sa riadi vzorcom 3: 1
23. Genotyp rodičovských foriem hrachu („čisté línie“) v Mendelových experimentoch
24. Genotyp potomkov (hybridy 1. generácie) u hrachu v Mendelových experimentoch
25. Hrachová farba prvej generácie
26. F1 Mendel sa dostal opeľovaním
27. F2 Mendel získaný opeľovaním
28. Typ štiepenia pri krížení heterozygotného a homozygotného pre recesívny
29. Monoheterozygotné formy ... odrody gamét
30. Hybridy s monohybridným krížením a úplnou dominanciou tvoria ... fenotypové triedy
31. Mendelove zákony sa týkali tohto druhu dedičstva
32. Nealelické gény umiestnené na rovnakom chromozóme, ak nedôjde k kríženiu, sú dedičné
33. Autor chromozomálnej teórie dedičnosti
34. Chromozómy, ktoré sa líšia veľkosťou a tvarom u mužov a žien
35. Chromozómy, ktoré sa nelíšia veľkosťou a tvarom
36. Nachádzajú sa gény „viazané na pohlavie“
37. Príkladom je hemofília a farebná slepota u ľudí a dedičnosť farby srsti u mačiek
38. Gény týchto vlastností sú lokalizované
39. Samce cicavcov a Drosophila sú heterozygotné kvôli znakom spojeným so sexom
40. Náhodné zmeny v genetickom materiáli jednotlivca
41. Zmena počtu chromozómov
42. Zmena jedného alebo viacerých nukleotidov
43. Zmeny v sekvencii aminokyselín bielkovín
44. Autor zákona o homológnych radoch dedičnej variability
45. Hlavná metóda genetiky, nepoužiteľná v ľudskej genetike
46. Metóda ľudskej genetiky, založená na farbení chromozómov a následnom skúmaní ich veľkosti a tvaru
47. Metóda ľudskej genetiky založená na analýze rodokmeňov a sledovaní prenosu určitého znaku
48. Metóda ľudskej genetiky založená na analýze fenotypových prejavov znakov u identických dvojčiat
49. Metóda ľudskej genetiky založená na analýze enzymatickej aktivity bielkovín, ktoré katalyzujú dôležité fyziologické procesy
50. Látky spôsobujúce mutácie
Úvod.
Genetika je veda, ktorá študuje zákony dedičnosti a variability živých organizmov.
Človek už dlho zaznamenal tri javy súvisiace s dedičnosťou: po prvé, podobnosť vlastností potomkov a rodičov; za druhé, rozdiely medzi niektorými (niekedy mnohými) znakmi potomstva a zodpovedajúcimi rodičovskými vlastnosťami; po tretie, výskyt postáv, ktoré boli iba u vzdialených predkov, u potomkov. Kontinuita znakov medzi generáciami je zaistená procesom hnojenia. Človek od nepamäti spontánne využíval vlastnosti dedičnosti na praktické účely - na šľachtenie odrôd pestovaných rastlín a plemien domácich zvierat.
Prvé predstavy o mechanizme dedičnosti vyjadrili starovekí grécki vedci Demokritos, Hippokrates, Platón, Aristoteles. Autor prvej vedeckej evolučnej teórie J.-B. Lamarck pomocou myšlienok starovekých gréckych vedcov vysvetlil, čo postuloval na prelome 18.-19. storočia. princíp prenosu nových vlastností získaných počas života jednotlivca na potomstvo. C. Darwin predložil teóriu pangenézy, ktorá vysvetľuje dedičnosť získaných znakov
Definoval Charles Darwin dedičnosť ako vlastnosť všetkých živých organizmov prenášať svoje vlastnosti a vlastnosti z generácie na generáciu, a variabilita ako vlastnosť všetkých živých organizmov získavať nové vlastnosti v procese individuálneho vývoja.
Vlastnosti sa dedia reprodukciou. Sexuálnou reprodukciou vznikajú v dôsledku oplodnenia nové generácie. Hmotné základy dedičnosti sú obsiahnuté v zárodočných bunkách. Pri asexuálnej alebo vegetatívnej reprodukcii sa nová generácia vyvíja buď z jednobunkových spór, alebo z mnohobunkových formácií. A pri týchto formách reprodukcie sa spojenie medzi generáciami uskutočňuje prostredníctvom buniek, ktoré obsahujú materiálne základy dedičnosti (elementárne jednotky dedičnosti) - gény - sú časti DNA chromozómov.
Súbor génov, ktoré organizmus dostáva od rodičov, tvorí jeho genotyp. Všetky vonkajšie a vnútorné vlastnosti sú fenotypom. Fenotyp sa vyvíja v dôsledku interakcie genotypu a podmienok prostredia. Tak či onak, základom sú črty, ktoré nesú v sebe gény.
Vzory, ktorými sa znaky prenášajú z generácie na generáciu, prvýkrát objavil veľký český vedec Gregor Mendel. Objavil a sformuloval tri zákony dedičnosti, ktoré tvorili základ modernej genetiky.
Život a vedecký výskum Gregora Johanna Mendela.
Moravský mních a genetik rastlín. Johann Mendel sa narodil v roku 1822 v meste Heinzendorf (dnes Gincice v Českej republike), kde jeho otec vlastnil malý roľnícky prídel. Gregor Mendel bol podľa svedectiev tých, ktorí ho poznali, skutočne milým a príjemným človekom. Po získaní základného vzdelania v miestnej dedinskej škole a neskôr, po absolvovaní kolégia piaristov v Leipnicku, bol v roku 1834 prijatý na prvú gramatickú triedu na cisársko-kráľovské gymnázium Troppaun. O štyri roky neskôr boli Johannini rodičia v dôsledku sútoku mnohých nešťastných udalostí, ktoré na seba rýchlo nasledovali, úplne zbavení možnosti uhradiť nevyhnutné výdavky spojené so vzdelaním a ich vtedy iba 16 -ročný syn mal nútený úplne nezávisle sa starať o svoju vlastnú údržbu ... V roku 1843 bol Mendel prijatý do augustiniánskeho kláštora svätého Tomáša v Altbrunne, kde prijal meno Gregor. V roku 1846 sa Mendel zúčastnil aj prednášok o poľnohospodárstve, záhradníctve a vinárstve na Filozofickom inštitúte v Brunne. V roku 1848, po absolvovaní kurzu teológie, dostal Mendel s hlbokým rešpektom povolenie študovať na doktorandských skúškach. Keď budúci rok posilnil svoj zámer na vyšetrenie, dostal príkaz zaujať miesto stúpenca cisársko-kráľovského gymnázia v Znaime, ktoré s radosťou nasledoval.
V roku 1851 poslal kláštorný opát Mendela na štúdium na viedenskú univerzitu, kde okrem iného študoval botaniku. Po skončení univerzity Mendel učil vedu na miestnej škole. Vďaka tomuto kroku sa jeho finančná situácia radikálne zmenila. V prospešnom blahu fyzickej existencie, tak potrebnom pre každé prenasledovanie, sa mu odvaha a sila vrátili s hlbokou úctou a počas skúšobného roku s veľkou usilovnosťou a láskou študoval predpísané klasické predmety. Vo voľných hodinách sa venoval malej botanickej a mineralogickej zbierke, ktorá mu bola v kláštore k dispozícii. Jeho vášeň pre oblasť prírodných vied bola tým väčšia, čím viac príležitostí dostával sa mu. Aj keď ten, ktorý bol spomenutý v týchto lekciách, bol zbavený akéhokoľvek vedenia a cesta autodidaktu tu, ako v žiadnej inej vede, je náročná a pomaly vedie k cieľu, napriek tomu počas tejto doby Mendel získal takú lásku k štúdiu prírody, že nešetril svojim úsilím.vyplniť svoje zmenené medzery samoštúdiom a podľa rád ľudí s praktickými skúsenosťami. 3. apríla 1851 sa „učiteľský zbor“ školy rozhodol pozvať pána Gregora Mendela, kanonika kláštora sv. Pomologické úspechy Gregora Mendela mu vyniesli právo na hviezdny titul a prechodné miesto priaznivca prírodopisu v prípravnej triede Technickej školy. V prvom semestri štúdia študoval iba desať hodín týždenne a iba s Dopplerom. V druhom semestri sa učil dvadsať hodín týždenne. Z toho desať - vo fyzike s Dopplerom, päť týždenne - v zoológii s Rudolfom Knerom. Jedenásť hodín týždenne - botanika u profesora Fenzla: okrem prednášok o morfológii a taxonómii absolvoval aj špeciálny workshop o popise a definícii rastlín. V treťom semestri sa prihlásil na tridsaťdva hodín výučby týždenne: desať hodín pre fyziku u Dopplera, desať pre chémiu u Rottenbachera: všeobecná chémia, lekárska chémia, farmakologická chémia a workshop analytickej chémie. Päť pre Knerovu zoológiu. Šesť hodín tréningu s Ungerom, jedným z prvých cytologov na svete. Vo svojich laboratóriách študoval anatómiu a fyziológiu rastlín a absolvoval workshop o mikroskopických technikách. A opäť - raz týždenne na Katedre matematiky - workshop o logaritme a trigonometrii.
1850, život išiel dobre. Mendel sa už dokázal udržať a tešil sa veľkému rešpektu svojich kolegov, pretože sa dobre vyrovnal so svojimi povinnosťami a veľmi príjemne sa s ním hovorilo. Jeho učeníci ho milovali.
V roku 1851 Gregor Mendel pristúpil k zásadnej otázke biológie - problému variability a dedičnosti. V tom čase začal vykonávať experimenty s cieleným pestovaním rastlín. Mendel priniesol rôzne rastliny z ďalekého i blízkeho okolia Brunnu. Rastliny pestoval v skupinách v časti kláštornej záhrady špeciálne určenej pre každú z nich za rôznych vonkajších podmienok. Zaoberal sa starostlivými meteorologickými pozorovaniami. Gregor väčšinu svojich experimentov a pozorovaní vykonával s hráškom, ktorý od roku 1854 každú jar každú jar vysieval do malej záhradky pod oknami prelatúry. Ukázalo sa, že je ľahké vytvoriť jasný hybridizačný experiment na hrachu. Aby ste to urobili, stačí otvoriť veľký, aj keď ešte nezrelý kvet, pinzetou, odrezať prašníky a nezávisle určiť „pár“ na kríženie. Pretože je samoopelenie vylúčené, odrody hrachu sú spravidla „čisté línie“ s konštantnými charakteristikami, ktoré sa nemenia z generácie na generáciu, ktoré sú veľmi jasne načrtnuté. Mendel vyzdvihol vlastnosti, ktoré určovali medziodvetvové rozdiely: farba šupky zrelých zŕn a oddelene od nezrelých zŕn, tvar zrelého hrachu, farba „proteínu“ (endospermu), dĺžka osi stonky , umiestnenie a farba púčikov. Na experiment použil viac ako tridsať odrôd a každá z odrôd bola predtým podrobená dvojročnému testu na „stálosť“, na „stálosť vlastností“, na „čistotu krvi“ - v roku 1854 a v roku 1855. Experimenty s hráškom pokračovali osem rokov. Stokrát v ôsmich kvetoch vlastnými rukami opatrne odrezal prašníky a napísaním peľu z tyčiniek kvetu inej odrody na pinzetu aplikoval na stigmu piestika. Pre desaťtisíc rastlín získaných krížením a zo samoopelených hybridov bolo zavedených desaťtisíc pasov. Záznamy v nich sú úhľadné: keď rodičovská rastlina rástla, aké mala kvety, ktorých peľ bol oplodnený, ktorý hrášok - žltý alebo zelený, hladký alebo vráskavý - bol získaný, ktoré kvety - farba na okrajoch, farba v stred - rozkvitol, keď boli prijaté semená, koľko z nich je žltých, koľko je zelených, okrúhlych, vráskavých, koľko z nich je vybraných na výsadbu, kedy sú zasadené a podobne.
Výsledkom jeho výskumu bola správa „Experimenty na rastlinných hybridoch“, ktorú v roku 1865 prečítal brunnský prírodovedec. V správe sa uvádza: „Dôvodom na založenie experimentov, ktorým je tento článok venovaný, bolo umelé kríženie okrasných rastlín, ktoré sa uskutočnilo s cieľom získať nové formy, ktoré sa líšia farbou. Založenie ďalších experimentov s cieľom sledovať vývoj hybridov u ich potomkov dalo impulz nápadnej pravidelnosti, s ktorou sa hybridné formy neustále vracali do svojich rodičovských foriem. Ako sa často stáva v dejinách vedy, Mendelovmu dielu sa okamžite nedostalo náležitého uznania od jeho súčasníkov. Výsledky jeho experimentov boli publikované na stretnutí Spoločnosti pre prírodné vedy mesta Brunn a potom publikované v časopise tejto spoločnosti, ale Mendelove myšlienky v tom čase nenašli podporu. Vydanie časopisu popisujúceho Mendelovo revolučné dielo zhromažďuje prach v knižniciach už tridsať rokov. Až na konci 19. storočia objavili vedci zaoberajúci sa problémami dedičnosti Mendelove diela a on mohol získať (už posmrtne) zaslúžené uznanie.
Mendelov prvý zákon
Kríženie dvoch organizmov sa nazýva hybridizácia, sa nazýva potomstvo krížením dvoch jedincov s rôznou dedičnosťou Hybrid, a samostatná osoba - hybrid.MonoHybrid sa nazýva kríženie dvoch organizmov, ktoré sa navzájom líšia v jednom páre alternatívnych (navzájom sa vylučujúcich) vlastností. V dôsledku toho sa pri takom krížení sledujú vzorce dedičnosti iba dvoch znakov, ktorých vývoj je spôsobený dvojicou alelických génov. Všetky ostatné znaky charakteristické pre tieto organizmy sa neberú do úvahy.
Ak krížite rastliny hrachu so žltými a zelenými semenami, potom všetky výsledné hybridy budú mať žlté semená. Rovnaký obraz je pozorovaný pri krížení rastlín s hladkým a vráskavým tvarom semien; všetci potomkovia prvej generácie budú mať hladký tvar semien:
Preto bol pomenovaný prvý Mendelov zákon Zákon uniformity hybridov prvej generácie.
Ak použijeme termíny, ktoré sa objavili mnoho rokov po Mendelovej práci, potom môžeme povedať, že bunky hrachových rastlín jednej odrody obsahujú dva gény iba žltej farby a gény rastlín inej odrody - dva gény iba zelenej farby. Nazývajú sa gény zodpovedné za vývoj jedného znaku (napríklad zafarbenia semien) alelické gény... V dôsledku toho sa v hybride prvej generácie vyvíja iba jeden z každého páru alternatívnych vlastností. Zdá sa, že druhé znamenie zmizne, neobjaví sa. Fenomén prevahy v hybridnom sznamenie jedného z rodičov G. Mendel pomenovaný dominprídel. Bol pomenovaný znak, ktorý sa prejavuje hybridom prvej generácie a potláča vývoj iného znaku dominantný a naopak, to znamená, že je potlačené, podpíše - recesívny. Gregor Mendel sa rozhodol nahradiť popis vlastností rastlín abstraktným kódom „A, B, C, D, E, F, G“ a „a, b, c, d, e, f, g“ a potom pozorovaním osud jedného páru vlastností prešiel k sledovaniu dvoch, troch, štyroch párov súčasne. Veľký A, B, C, D, E, F, G, vyznačil dominanty; malé a, b, c, d, e, f, g - recesívne. Ak existujú dva identické alelické gény v genotype organizmu (zygota), oba sú dominantné alebo oba sú recesívne (AA alebo aa), takýto organizmus sa nazýva homozygotný. Ak je z páru alelických génov jeden dominantný a druhý recesívny (Aa), vtedy sa taký organizmus nazýva heterozygotní.
Štiepny zákon, alebo druhý Mendelov zákon.
Ak sa potomkovia prvej generácie, identickí v skúmanom znaku, navzájom skrížia, potom sa v druhej generácii znaky oboch rodičov prejavia v určitom číselnom pomere: 3/4 jedincov bude mať dominantný znak, ¼ recesívny:

Fenomén, v ktorom dochádza k kríženiu heterozygotnýchjednotlivci vedú k tvorbe potomkov, z ktorých niektorírogo nesie dominantu, a časť- recesívnyny, sa nazýva rozdelenie. V dôsledku toho recesívny znak u hybridov prvej generácie nezmizol, bol iba potlačený a prejaví sa v druhej hybridnej generácii.
Hypotéza čistoty gamét. Mendel naznačil, že počas tvorby hybridov sa dedičné faktory nemiešajú, ale zostávajú nezmenené. V hybride sú prítomné oba faktory - dominantný a recesívny, ale dominantný dedičný faktor sa prejavuje formou znaku, pričom recesívny faktor je potlačený. Komunikácia medzi generáciami počas sexuálnej reprodukcie sa uskutočňuje prostredníctvom pohlavných buniek - hameta. Preto treba predpokladať, že každá gaméta nesie z páru iba jeden faktor. Potom počas hnojenia fúzia dvoch gamét, z ktorých každá nesie recesívny dedičný faktor, povedie k vytvoreniu organizmu s recesívnym znakom, ktorý sa prejavuje fenotypicky. Fúzia gamét, z ktorých každá nesie dominantný faktor, alebo dvoch gamét, z ktorých jedna obsahuje dominantný faktor a druhá recesívny faktor, povedie k vývoju organizmu s dominantnou črtou.
Mendel vysvetlil rozdelenie potomstva pri krížení heterozygotných jedincov tým, že gaméty sú geneticky čisté, to znamená, že nesú iba jeden gén z alelického páru. Hypotéza(teraz sa tomu hovorí zákon) čistotagaméty možno formulovať nasledovne: počas tvorby zárodočných buniek sa do každej gaméty dostane iba jeden gén z alelického páru.
Prečo a ako sa to deje? Je známe, že každá bunka tela má presne rovnakú diploidnú sadu chromozómov. Dva homológne chromozómy obsahujú dva identické gény. Geneticky „čisté“ gaméty sa tvoria nasledovne: keď sa zlúčia samčie a samičie gaméty, získa sa hybrid s diploidnou (dvojitou) sadou chromozómov.
Ako je zrejmé z diagramu (dodatok 2), zygota prijíma polovicu chromozómov z otcovského organizmu a polovicu z materského.
V procese tvorby gamét v hybride sa do rôznych buniek dostávajú aj homológne chromozómy počas prvého meiotického delenia.
Pre tento alelický pár sa tvoria dve odrody gamét. Pri oplodnení sa náhodne stretnú gaméty nesúce rovnaké alebo rôzne alely. Vzhľadom na štatistickú pravdepodobnosť, s dostatočne veľkým počtom gamét u potomstva, bude 25% genotypov homozygotne dominantných, 50% - heterozygotných, 25% - homozygotných recesívnych, t.j. pomer 1AA: 2Aa: 1aa.
Podľa fenotypu je potomstvo druhej generácie počas monohybridného kríženia distribuované v pomere 3: 1 (¾ jedincov s dominantným znakom, ¼ jedincov s recesívnym znakom).
Cytologickým základom štiepenia znakov pri monohybridnom krížení je divergencia homológnych chromozómov k rôznym pólom bunky a tvorba haploidných zárodočných buniek pri meióze.
Vo vyššie uvedených príkladoch bolo pravidlo jednotnosti vyjadrené v skutočnosti, že všetky hybridy boli navonok podobné jednému z rodičov. Nie vždy to tak je. Znaky v heterozygotných formách majú často prechodný charakter, t.j. dominancia nemusí byť úplná. Schéma kríženia dvoch dedičných foriem rastliny Night Beauty:


Jeden z nich má červené kvety (a to je dominanta) a druhý je biely. Diagram ukazuje, že všetky hybridy prvej generácie majú ružové kvety. V druhej generácii dochádza k štiepeniu v pomere 1: 2: 1, t.j. jeden červený kvet (homozygot), dva ružové kvety (heterozygot), jeden biely (homozygot). Tento jav sa nazýva neúplná dominancia.
V prípade neúplnej dominancie dominantný gén v heterozygotnom stave nie vždy úplne potlačí recesívny gén. V niektorých prípadoch hybrid fi nereprodukuje úplne žiadny z rodičovských znakov a tento znak je strednej povahy s viac -menej odchýlkou od dominantného alebo recesívneho stavu. Ale všetci jednotlivci tejto generácie sú v tejto charakteristike jednotní. Rozšírená je neúplná dominancia. Zistilo sa to pri štúdiu dedičnosti farby kvetov u snapdragons, farby vlny u hovädzieho dobytka a oviec, biochemických vlastností u ľudí atď. Stredné znaky vyplývajúce z neúplnej dominancie často predstavujú pre ľudí estetickú alebo materiálnu hodnotu. Vzniká otázka: je možné chovať výberom napríklad odrodu nočnej krásky s ružovou farbou kvetov? Očividne nie, pretože táto vlastnosť sa vyvíja iba u heterozygotov a keď sú navzájom skrížení, vždy dôjde k rozdeleniu:
Rozšírená je neúplná dominancia. Zistilo sa to pri štúdiu dedičnosti farby kvetov u snapdragons, farby vlny u hovädzieho dobytka a oviec, biochemických vlastností u ľudí atď. Stredné znaky vyplývajúce z neúplnej dominancie často predstavujú pre ľudí estetickú alebo materiálnu hodnotu. Vzniká otázka: je možné chovať výberom napríklad odrodu nočnej krásky s ružovou farbou kvetov? Očividne nie, pretože táto vlastnosť sa vyvíja iba u heterozygotov a keď sú navzájom skrížení, vždy dôjde k rozdeleniu.
Zákon nezávislej kombinácie alebo tretíMendelov zákon.
Mendelova štúdia dedičnosti jedného páru alel umožnila stanoviť množstvo dôležitých genetických vzorcov: fenomén dominancie, nemennosť recesívnych alel u hybridov, štiepenie potomkov hybridov v pomere 3: 1, a tiež predpokladať, že gaméty sú geneticky čisté, to znamená, že obsahujú iba jeden gén z alelických párov. Organizmy sa však líšia v mnohých génoch. Je možné stanoviť vzorce dedičnosti dvoch párov alternatívnych znakov a ďalších pomocou dihybridný alebo polyhybridné kríženie, t.j. kríženie rodičovských foriem líšiacich sa dvoma pármi znakov.
Na dihybridné kríženie vzal Mendel homozygotné hrachové rastliny, ktoré sa líšia dvoma ukazovateľmi - farbou semena (žltá, zelená) a tvarom semena (hladký, vráskavý). Dominantné vlastnosti - žlté sfarbenie (A) a hladký tvar (V) semená. Každá rastlina tvorí pre študované alely jednu škálu gamét: Keď sa gaméty zlúčia, všetky potomstvo bude jednotné:


|
Pannetová mriežka |
||||||

Je ľahké vypočítať, že podľa fenotypu sa potomstvo delí na 4 skupiny: 9 žltých hladkých, 3 žltých vráskavých, 3 zelených hladkých, 1 žltých vráskavých (9: 3: 3: 1). Ak vezmeme do úvahy výsledky štiepenia pre každý pár znakov zvlášť, ukazuje sa, že pomer počtu žltých semien k počtu zelených semien a pomeru hladkých a vráskavých semien pre každý pár je 3: 1. To možno vyjadriť algebraicky ako štvorec binomického čísla
(3 + 1) ² = 3² + 2 · 3 + 1² alebo 9 + 3 + 3 + 1
Pri dihybridnom krížení sa teda každý pár znakov počas delenia u potomkov správa rovnako ako pri monohybridnom krížení, to znamená nezávisle od druhého páru znakov.
Pri oplodnení sa gaméty kombinujú podľa pravidiel náhodných kombinácií, ale s rovnakou pravdepodobnosťou pre každú z nich. Vo výsledných zygotoch vznikajú rôzne kombinácie génov. Teraz môžeme sformulovať tretí Mendelov zákon: pri krížení dvoch homozygotných jedincov, odlišnýchoddelené od seba navzájom dva alebo viac párov alternatívnych znakov, génov aich zodpovedajúce vlastnosti sú zdedené nezávisleod seba navzájom a sú kombinované vo všetkých možnýchkombinácie.
Mendelove zákony slúžia ako základ pre analýzu rozdelenia v zložitejších prípadoch: keď sa jednotlivci líšia v troch, štyroch pároch znakov alebo vo viacerých.
Podmienky súladu s Mendelovými dedičskými zákonmi
Zákony objavené Gregorom Mendelom nie sú vždy použiteľné v genetike. Podmienok dodržiavania Mendelových zákonov je mnoho. Pre takéto prípady existujú ďalšie zákony (napríklad: Morganov zákon) alebo vysvetlenia.
Sformulujme základné podmienky dodržiavania dedičských zákonov.
Aby bol dodržaný zákon jednotnosti hybridov prvej generácie, je potrebné, aby:
rodičovské organizmy boli homozygotné;
gény rôznych alel boli umiestnené v rôznych chromozómoch, a nie v jednom (v opačnom prípade môže dôjsť k javu „prepojenej dedičnosti“).
Zákon o štiepení bude dodržaný, ak
u hybridov zostávajú dedičné faktory nezmenené;
Zákon nezávislej distribúcie génov u potomkov a vznik rôznych kombinácií týchto génov počas dihybridného kríženia je možný iba v prípade.
ak sa páry alelických génov nachádzajú v rôznych pároch homológnych chromozómov.
Porušenie týchto podmienok môže viesť buď k absencii rozdelenia v druhej generácii, alebo k rozdeleniu v prvej generácii; alebo k skresleniu pomeru rôznych genotypov a fenotypov. Mendelove zákony sú univerzálne pre všetky pohlavne sa rozmnožujúce diploidné organizmy. Vo všeobecnosti sú platné pre autozomálne gény s plnou penetráciou (100% frekvencia prejavu analyzovaného znaku; 100% penetrácia znamená, že znak je vyjadrený vo všetkých nosičoch alely, ktorá určuje vývoj tohto znaku) a konštantnou expresivitou; konštantná expresivita znamená, že fenotypová závažnosť znaku je rovnaká alebo približne rovnaká vo všetkých nosičoch alely, ktorá určuje vývoj tohto znaku.
Záver.
Zákony Gregora Mendela sú v súčasnosti široko používané v šľachtiteľských rastlinách, zvieratách a mikroorganizmoch, v medicíne, genetickom inžinierstve a mnohých ďalších odvetviach ľudského života, ako aj pri riešení problémov v genetike.
Je dôležité poznamenať, že Mendel formuloval zákony a vyvodil závery v čase, keď nebolo nič známe o DNA, génoch a chromozómoch. Ukázalo sa však, že má úplnú pravdu, a aj keď nie okamžite, jeho teórie boli uznané a brané ako základ rozvíjajúcej sa vedy o genetike.
Mendelovská teória dedičnosti, t.j. súhrn myšlienok o dedičných determinantoch a povahe ich prenosu z rodičov na potomkov vo svojom zmysle je v priamom protiklade s domainelskými teóriami, najmä s Darwinovou teóriou pangenézy. V súlade s touto teóriou sú znaky rodičov priame, t.j. zo všetkých častí tela, prenesený na potomstvo. Charakter znaku potomka by preto mal priamo závisieť od vlastností rodiča. To je úplne v rozpore so závermi, ktoré urobil Mendel: determinanty dedičnosti, t.j. gény sú v tele prítomné relatívne nezávisle na sebe. Charakter znakov (fenotyp) je určený ich náhodnou kombináciou. Nie sú upravené žiadnou časťou tela a sú vo vzťahu dominancie a recesie. Mendelovská teória dedičnosti je teda proti myšlienke dedičnosti postáv získaných počas individuálneho vývoja.
Mendelove experimenty slúžili ako základ pre rozvoj modernej genetiky - vedy, ktorá študuje dve hlavné vlastnosti organizmu - dedičnosť a variabilitu. Podarilo sa mu identifikovať vzorce dedičnosti vďaka zásadne novým metodologickým prístupom:
1) Mendel si úspešne vybral predmet výskumu;
2) analyzoval dedičnosť jednotlivých znakov u potomkov krížených rastlín, ktoré sa líšia v jednom, dvoch a troch pároch kontrastných alternatívnych znakov. V každej generácii bol vedený záznam oddelene pre každý pár týchto charakteristík;
3) nielen zaznamenal získané výsledky, ale tiež vykonal ich matematické spracovanie.
Uvedené jednoduché metódy výskumu predstavovali zásadne novú, hybridologickú metódu štúdia dedičnosti, ktorá sa stala základom pre ďalší výskum genetiky.
Bibliografia.
Všeobecná biológia: Učebnica pre 9-10 ročníkov. Streda škola / Polyansky Yu.I., Brown A.D., Verzilin N.M. a ďalší; M.: Education, 1987.-287 s.: Chorý.
Mendel dospel k záveru, že predispozícia k pôrodu ... ovplyvňuje frekvencie genotypov v populácii. Zákony Mendel nehovorte nám nič o frekvenciách ...Mendelov prvý zákon
Kríženie dvoch organizmov sa nazýva hybridizácia, sa nazýva potomstvo krížením dvoch jedincov s rôznou dedičnosťou Hybrid, a samostatná osoba - hybrid. Mono Hybrid sa nazýva kríženie dvoch organizmov, ktoré sa navzájom líšia v jednom páre alternatívnych (navzájom sa vylučujúcich) vlastností. V dôsledku toho sa pri takom krížení sledujú vzorce dedičnosti iba dvoch znakov, ktorých vývoj je spôsobený dvojicou alelických génov. Všetky ostatné znaky charakteristické pre tieto organizmy sa neberú do úvahy.
Ak krížite rastliny hrachu so žltými a zelenými semenami, potom všetky výsledné hybridy budú mať žlté semená. Rovnaký obraz je pozorovaný pri krížení rastlín s hladkým a vráskavým tvarom semien; všetci potomkovia prvej generácie budú mať hladký tvar semien:
Preto bol pomenovaný prvý Mendelov zákon Zákon uniformity hybridov prvej generácie.
Ak použijeme termíny, ktoré sa objavili mnoho rokov po Mendelovej práci, potom môžeme povedať, že bunky hrachových rastlín jednej odrody obsahujú dva gény iba žltej farby a gény rastlín inej odrody - dva gény iba zelenej farby. Nazývajú sa gény zodpovedné za vývoj jedného znaku (napríklad zafarbenia semien) alelické gény... V dôsledku toho sa v hybride prvej generácie vyvíja iba jeden z každého páru alternatívnych vlastností. Zdá sa, že druhé znamenie zmizne, neobjaví sa. Fenomén prevahy v hybridnom s znamenie jedného z rodičov G. Mendel pomenovaný domin prídel. Bol pomenovaný znak, ktorý sa prejavuje hybridom prvej generácie a potláča vývoj iného znaku dominantný a naopak, to znamená, že je potlačené, podpíše - recesívny. Gregor Mendel sa rozhodol nahradiť popis vlastností rastlín abstraktným kódom „A, B, C, D, E, F, G“ a „a, b, c, d, e, f, g“ a potom pozorovaním osud jedného páru vlastností prešiel k sledovaniu dvoch, troch, štyroch párov súčasne. Veľký A, B, C, D, E, F, G, vyznačil dominanty; malé a, b, c, d, e, f, g - recesívne. Ak existujú dva identické alelické gény v genotype organizmu (zygota), oba sú dominantné alebo oba sú recesívne (AA alebo aa), takýto organizmus sa nazýva homozygotný. Ak je z páru alelických génov jeden dominantný a druhý recesívny (Aa), vtedy sa taký organizmus nazýva heterozygotní.
Štiepny zákon, alebo druhý Mendelov zákon.
Ak sa potomkovia prvej generácie, identickí v skúmanom znaku, navzájom skrížia, potom sa v druhej generácii znaky oboch rodičov prejavia v určitom číselnom pomere: 3/4 jedincov bude mať dominantný znak, ¼ recesívny:

Fenomén, v ktorom dochádza k kríženiu heterozygotných jednotlivci vedú k tvorbe potomkov, z ktorých niektorí rogo nesie dominantu, a časť - recesívny ny, sa nazýva rozdelenie. V dôsledku toho recesívny znak u hybridov prvej generácie nezmizol, bol iba potlačený a prejaví sa v druhej hybridnej generácii.
Hypotéza čistoty gamét. Mendel naznačil, že počas tvorby hybridov sa dedičné faktory nemiešajú, ale zostávajú nezmenené. V hybride sú prítomné oba faktory - dominantný a recesívny, ale dominantný dedičný faktor sa prejavuje formou znaku, pričom recesívny faktor je potlačený. Komunikácia medzi generáciami počas sexuálnej reprodukcie sa uskutočňuje prostredníctvom pohlavných buniek - ha meta. Preto treba predpokladať, že každá gaméta nesie z páru iba jeden faktor. Potom počas hnojenia fúzia dvoch gamét, z ktorých každá nesie recesívny dedičný faktor, povedie k vytvoreniu organizmu s recesívnym znakom, ktorý sa prejavuje fenotypicky. Fúzia gamét, z ktorých každá nesie dominantný faktor, alebo dvoch gamét, z ktorých jedna obsahuje dominantný faktor a druhá recesívny faktor, povedie k vývoju organizmu s dominantnou črtou.
Mendel vysvetlil rozdelenie potomstva pri krížení heterozygotných jedincov tým, že gaméty sú geneticky čisté, to znamená, že nesú iba jeden gén z alelického páru. Hypotéza(teraz sa tomu hovorí zákon) čistota gaméty možno formulovať nasledovne: počas tvorby zárodočných buniek sa do každej gaméty dostane iba jeden gén z alelického páru.
Prečo a ako sa to deje? Je známe, že každá bunka tela má presne rovnakú diploidnú sadu chromozómov. Dva homológne chromozómy obsahujú dva identické gény. Geneticky „čisté“ gaméty sa tvoria nasledovne: keď sa zlúčia samčie a samičie gaméty, získa sa hybrid s diploidnou (dvojitou) sadou chromozómov.
Ako je zrejmé z diagramu (dodatok 2), zygota prijíma polovicu chromozómov z otcovského organizmu a polovicu z materského.
V procese tvorby gamét v hybride sa do rôznych buniek dostávajú aj homológne chromozómy počas prvého meiotického delenia.
Pre tento alelický pár sa tvoria dve odrody gamét. Pri oplodnení sa náhodne stretnú gaméty nesúce rovnaké alebo rôzne alely. Vzhľadom na štatistickú pravdepodobnosť, s dostatočne veľkým počtom gamét u potomstva, bude 25% genotypov homozygotne dominantných, 50% - heterozygotných, 25% - homozygotných recesívnych, t.j. pomer 1AA: 2Aa: 1aa.
Podľa fenotypu je potomstvo druhej generácie počas monohybridného kríženia distribuované v pomere 3: 1 (¾ jedincov s dominantným znakom, ¼ jedincov s recesívnym znakom).
Cytologickým základom štiepenia znakov pri monohybridnom krížení je divergencia homológnych chromozómov k rôznym pólom bunky a tvorba haploidných zárodočných buniek pri meióze.
Vo vyššie uvedených príkladoch bolo pravidlo jednotnosti vyjadrené v skutočnosti, že všetky hybridy boli navonok podobné jednému z rodičov. Nie vždy to tak je. Znaky v heterozygotných formách majú často prechodný charakter, t.j. dominancia nemusí byť úplná. Schéma kríženia dvoch dedičných foriem rastliny Night Beauty:


Jeden z nich má červené kvety (a to je dominanta) a druhý je biely. Diagram ukazuje, že všetky hybridy prvej generácie majú ružové kvety. V druhej generácii dochádza k štiepeniu v pomere 1: 2: 1, t.j. jeden červený kvet (homozygot), dva ružové kvety (heterozygot), jeden biely (homozygot). Tento jav sa nazýva neúplná dominancia.
V prípade neúplnej dominancie dominantný gén v heterozygotnom stave nie vždy úplne potlačí recesívny gén. V niektorých prípadoch hybrid fi nereprodukuje úplne žiadny z rodičovských znakov a tento znak je strednej povahy s viac -menej odchýlkou od dominantného alebo recesívneho stavu. Ale všetci jednotlivci tejto generácie sú v tejto charakteristike jednotní. Rozšírená je neúplná dominancia. Zistilo sa to pri štúdiu dedičnosti farby kvetov u snapdragons, farby vlny u hovädzieho dobytka a oviec, biochemických vlastností u ľudí atď. Stredné znaky vyplývajúce z neúplnej dominancie často predstavujú pre ľudí estetickú alebo materiálnu hodnotu. Vzniká otázka: je možné chovať výberom napríklad odrodu nočnej krásky s ružovou farbou kvetov? Očividne nie, pretože táto vlastnosť sa vyvíja iba u heterozygotov a keď sú navzájom skrížení, vždy dôjde k rozdeleniu:
Rozšírená je neúplná dominancia. Zistilo sa to pri štúdiu dedičnosti farby kvetov u snapdragons, farby vlny u hovädzieho dobytka a oviec, biochemických vlastností u ľudí atď. Stredné znaky vyplývajúce z neúplnej dominancie často predstavujú pre ľudí estetickú alebo materiálnu hodnotu. Vzniká otázka: je možné chovať výberom napríklad odrodu nočnej krásky s ružovou farbou kvetov? Očividne nie, pretože táto vlastnosť sa vyvíja iba u heterozygotov a keď sú navzájom skrížení, vždy dôjde k rozdeleniu.
Zákon nezávislej kombinácie alebo tretí Mendelov zákon.
Mendelova štúdia dedičnosti jedného páru alel umožnila stanoviť množstvo dôležitých genetických vzorcov: fenomén dominancie, nemennosť recesívnych alel u hybridov, štiepenie potomkov hybridov v pomere 3: 1, a tiež predpokladať, že gaméty sú geneticky čisté, to znamená, že obsahujú iba jeden gén z alelických párov. Organizmy sa však líšia v mnohých génoch. Je možné stanoviť vzorce dedičnosti dvoch párov alternatívnych znakov a ďalších pomocou dihybridný alebo polyhybridné kríženie, t.j. kríženie rodičovských foriem líšiacich sa dvoma pármi znakov.
Vedec z Českej republiky Gregor Mendel (1822-1884) v rámci svojho výskumu kríženia rôznych odrôd hrachu odhalil určité vzorce, ktoré sú dnes široko známe pod názvom troch Mendelových zákonov.
Prvý zákon
Ak boli rastlinné vzorky získané v prvej generácii krížením homozygotných rodičovských odrôd (čisté línie) s rôznymi znakmi, potomstvo je v genotype a fenotype rovnaké. V tomto prípade sa znakom rozumie akákoľvek vybraná kvalita rastlín, podľa ktorej je možné rozlíšiť dve odrody. Mendelov prvý zákon je tiež známy ako pravidlo dominancie alebo zákon uniformity pre hybridy prvej generácie. Znak, ktorý bol identifikovaný v prvej generácii, sa nazýval dominantný a kvalita, ktorá bola potláčaná a neprejavovala sa, bola recesívna.
Druhý zákon
Štiepny zákon. Jeho podstata spočíva v tom, že pri krížení, napríklad počas samoopelenia, rovnakého druhu hybridov prvej generácie v ďalšej generácii sa potomstvo delí podľa zvoleného znaku (fenotypu) v pomere 3 : 1 v prípade úplnej dominancie a 1: 2: 1 v prípade neúplnej dominancie.
Tretí zákon
Tretí Mendelov zákon sa zaoberá nezávislou dedičnosťou znakov alebo nezávislou kombináciou rastlinných génov. Ak dôjde k kríženiu rastlinných vzoriek, ktoré sa navzájom líšia v dvoch alebo viacerých pároch alternatívnych znakov, genetický materiál a fenotypové znaky, ktoré určujú, sú zdedené nezávisle od seba a môžu byť kombinované v akejkoľvek kombinácii.
Experimenty českého vedca sa stali základom pre rozvoj modernej genetiky. G. Mendel dokázal odhaliť základné vzorce dedičnosti pomocou najnovších metodických prístupov, ktoré stále používajú všetci genetici.
Pri jeho experimentoch všetky skrížené exempláre patrili k rovnakým rastlinným druhom. Je dôležité, aby sa zároveň navzájom zreteľne líšili v jednom, dvoch, niekoľkých pároch alternatívnych (opačných) znakov. Tieto vlastnosti boli konštantné, to znamená, že sa prenášali z generácie na generáciu pri krížení v rámci rovnakého druhu. Bolo potrebné študovať potomstvo z každého páru hybridov jednotlivo a urobiť kvantitatívny popis hybridov, ktoré sa líšia v určitých pároch alternatívnych znakov.
Tieto metódy výskumu tvorili základ novej hybridologickej metódy, s nástupom ktorej sa začala éra štúdia dedičnosti a variability.
Vzory dedičnosti znakov počas sexuálnej reprodukcie stanovil G. Mendel. Je potrebné jasne porozumieť genotypu a fenotypu, alelám, homo- a heterozygotnosti, dominancii a jej typom, typom krížov a tiež vypracovať schémy.
Monohybrid Hovorí sa o krížení, v ktorom sa rodičovské formy navzájom líšia v jednom páre kontrastných, alternatívnych znakov.
V dôsledku toho sa pri takom krížení sledujú vzorce dedičnosti iba dvoch variantov znaku, ktorých vývoj je spôsobený dvojicou alelických génov. Príkladmi monohybridných krížov vedených G. Mendelom sú hrachové kríže s takými jasne viditeľnými alternatívnymi znakmi, ako sú purpurové a biele kvety, žltá a zelená farba nezrelých plodov (fazuľa), hladký a vráskavý povrch semien, žltá a zelená farba atď.
Jednotnosť hybridov prvej generácie (Mendelov prvý zákon) Pri krížení hrachu s purpurovými (AA) a bielymi (aa) kvetmi Mendel zistil, že všetky hybridné rastliny prvej generácie (F1) mali purpurové kvety (obrázok 2).
Obrázok 2 – Schéma monohybridného kríženia
Súčasne sa neobjavila biela farba kvetu. Pri krížení rastlín, ktoré majú hladké a vráskavé semená, budú mať kríženci hladké semená. G. Mendel tiež zistil, že všetky hybridy F 1 boli jednotné (homogénne) pre každý zo siedmich znakov, ktoré študoval. V dôsledku toho sa u hybridov prvej generácie z páru alternatívnych vlastností rodičov objaví iba jeden a zdá sa, že znak druhého rodiča zmizne.
Alternatívne znaky sú navzájom sa vylučujúce a kontrastné znaky.
Mendel označil prevahu vlastností jedného z rodičov v hybridoch F1 za dominanciu a zodpovedajúcu vlastnosť za dominantnú. Znaky, ktoré sa u hybridov F 1 neprejavujú, označil za recesívne. Pretože všetky hybridy prvej generácie sú jednotné, tento jav sa nazýva Mendelov prvý zákon alebo hybridný zákon uniformity prvej generácie a pravidlo dominancie.
Možno to formulovať takto: pri krížení dvoch organizmov patriacich do rôznych čistých línií (dvoch homozygotných organizmov), ktoré sa navzájom líšia v jednom páre alternatívnych znakov, bude celá prvá generácia hybridov jednotná a bude niesť znak jedného z nich rodičia.
Každý gén má dva stavy - „A“ a „a“, takže tvoria jeden pár a každý člen páru sa nazýva alela. Gény nachádzajúce sa v rovnakých lokusoch (sekciách) homológnych chromozómov a určujúce alternatívny vývoj rovnakého znaku sa nazývajú alelické.
Napríklad purpurová a biela farba hrachového kvetu je dominantná a recesívna pre dve alely (A a a) jedného génu. Vzhľadom na prítomnosť dvoch alel sú možné dva stavy organizmu: homo- a heterozygotné. Ak organizmus obsahuje rovnaké alely konkrétneho génu (AA alebo aa), nazýva sa pre tento gén (alebo vlastnosť) homozygotný, a ak je odlišný (Aa), potom heterozygotný. Alela je preto formou existencie génu. Príkladom trialelického génu je gén, ktorý určuje systém krvných skupín ABO u ľudí. Existuje aj viac alel: pre gén, ktorý riadi syntézu ľudského hemoglobínu, ich je niekoľko desiatok.
Z hybridných semien hrachu Mendel vypestoval rastliny, ktoré podrobil samoopeleniu a výsledné semená znova vysiali. Výsledkom bola druhá generácia hybridov alebo hybridov F 2. Medzi nimi bolo rozdelenie zistené pre každý pár alternatívnych znakov v pomere asi 3: 1, tj tri štvrtiny rastlín mali dominantné znaky (fialové kvety, žlté semená, hladké semená atď.) A jedna štvrtina bola recesívne (biele kvety, zelené semená, vráskavé semená atď.). V dôsledku toho recesívny znak v hybride F1 nezmizol, ale bol iba potlačený a znova sa objavil v druhej generácii. Toto zovšeobecnenie sa neskôr nazývalo druhý Mendelov zákon alebo zákon rozdelenia.
Rozdelenie je jav, pri ktorom kríženie heterozygotných jedincov vedie k vzniku potomstva, z ktorých niektoré sú dominantné a niektoré sú recesívne.
Mendelov druhý zákon: keď sa v druhej generácii skrížia dvaja potomkovia prvej generácie (dvaja heterozygotní jedinci), rozdelenie sa pozoruje v určitom číselnom pomere: podľa fenotypu 3: 1, podľa genotypu 1: 2 : 1 (obr. 3).

Obrázok 3 - Schéma deliacich znakov
pri krížení krížencov F 1
Štiepenie znakov u potomkov pri krížení heterozygotných jedincov vysvetlil G. Mendel tým, že gaméty sú geneticky čisté, to znamená, že nesú iba jeden gén z alelického páru. Zákon čistoty gamét možno formulovať nasledovne: počas tvorby zárodočných buniek sa do každej gamety dostane iba jeden gén z alelického páru.
Je potrebné mať na pamäti, že použitie hybridologickej metódy na analýzu dedičnosti vlastností u akéhokoľvek druhu zvierat alebo rastlín zahŕňa nasledujúce krížiky:
kríženie rodičovských foriem (P), líšiace sa jedným (monohybridné kríženie) alebo niekoľkými pármi (polyhybridné kríženie) alternatívnych znakov a získaním hybridov prvej generácie (F 1);
kríženie hybridov F 1 navzájom a získanie hybridov druhej generácie (F 2);
matematická analýza výsledkov kríženia.
Následne sa Mendel obrátil na štúdiu dihybridného kríženia.
Dihybridný prechod Ide o kríženie, do ktorého sú zapojené dva páry alel (párové gény sú alelické a nachádzajú sa iba v homológnych chromozómoch).
Počas dihybridného kríženia G. Mendel študoval dedičnosť znakov, za ktoré sú zodpovedné gény ležiace v rôznych pároch homológnych chromozómov. V tomto ohľade musí každá gaméta obsahovať jeden gén z každého alelického páru.
Hybridy heterozygotné pre dva gény sa nazývajú diheterozygotné a ak sa líšia v troch alebo viacerých génoch, sú tri- a polyheterozygotné.
Zložitejšie schémy dihybridných krížov, zaznamenávanie genotypov a fenotypov F 2 sa vykonávajú pomocou Pennettovej mriežky. Uvažujme o príklade takého kríža. Na kríženie boli odobraté dve počiatočné homozygotné rodičovské formy: prvá forma mala žlté a hladké semená; druhá forma mala zelené a vráskavé semená (obr. 4).

Obrázok 4 - Dihybridné kríženie rastlín hrachu,
rôzneho tvaru a farby semien
Dominantnými znakmi sú žltá farba a hladké semená; zelená farba a vráskavé semená sú recesívne symptómy. Hybridy prvej generácie - krížené navzájom. V druhej generácii bolo fenotypové štiepenie pozorované v pomere 9: 3: 3: 1, alebo (3 + 1) 2, po samoopelení hybridov F1 v súlade so zákonom o štiepení sa znova objavili vráskavé a zelené semená.
Pôvodné rastliny v tomto prípade majú genotypy AABB a aabb a genotyp hybridov F 1 - AaBb, to znamená, že sú diheterozygotné.
Pri krížení heterozygotných jedincov líšiacich sa niekoľkými pármi alternatívnych znakov sa teda u potomstva pozoruje štiepenie fenotypu v pomere (3 + 1) n, kde n je počet párov alternatívnych znakov.
Gény, ktoré určujú vývoj rôznych párov znakov, sa nazývajú nealelické.
Výsledky kríženia dihybridných a polyhybridných závisia od toho, či sa gény, ktoré určujú uvažované znaky, nachádzajú v rovnakých alebo v rôznych chromozómoch. Mendel narazil na vlastnosti, ktorých gény boli v rôznych pároch homológnych chromozómov hrachu.
Pri meióze sú homológne chromozómy rôznych párov náhodne kombinované v gamétach. Ak sa otcovský chromozóm prvého páru dostane do gamety, potom sa s rovnakou pravdepodobnosťou do tejto gamety môžu dostať otcovský aj materský chromozóm druhého páru. Preto vlastnosti, ktorých gény sú v rôznych pároch homológnych chromozómov, sú kombinované nezávisle na sebe. Následne sa ukázalo, že zo siedmich párov znakov študovaných Mendelom na hrachu, v ktorých je diploidný počet chromozómov 2 n = 14, boli gény zodpovedné za jeden z párov znakov na tom istom chromozóme. Mendel však nenašiel porušenie zákona o nezávislej dedičnosti, pretože väzba medzi týmito génmi nebola pozorovaná kvôli veľkej vzdialenosti medzi nimi).
Na základe výskumu Mendel vyvodil tretí zákon - zákon nezávislej dedičnosti znakov alebo nezávislú kombináciu génov.
Každý pár alelických génov (a nimi riadených alternatívnych znakov) je zdedený nezávisle na sebe.
Zákon nezávislého kombinovania génov tvorí základ kombinačnej variability pozorovanej počas kríženia vo všetkých živých organizmoch. Poznamenávame tiež, že na rozdiel od prvého Mendelovho zákona, ktorý vždy platí, druhý zákon platí iba pre gény lokalizované v rôznych pároch homológnych chromozómov. Je to spôsobené tým, že nehomológne chromozómy sú v bunke kombinované nezávisle od seba, čo bolo dokázané nielen štúdiom charakteru dedičnosti znakov, ale aj priamou cytologickou metódou.
Pri štúdiu materiálu dávajte pozor na prípady porušení pravidelného štiepenia fenotypom spôsobených smrteľným pôsobením jednotlivých génov.
Dedičnosť a variabilita. Dedičnosť a variabilita sú najdôležitejšie vlastnosti charakteristické pre všetky živé organizmy.
Dedičná alebo genotypová variabilita je rozdelená na kombinačné a mutačné.
Variabilita sa nazýva kombinačná, ktorá je založená na tvorbe rekombinácií, to znamená na takých kombináciách génov, ktoré rodičia nemali.
V srdci kombinačnej variability je sexuálna reprodukcia organizmov, v dôsledku ktorej vzniká obrovská rozmanitosť genotypov. Tri procesy sú prakticky neobmedzenými zdrojmi genetických variácií:
Nezávislá divergencia homológnych chromozómov v prvom meiotickom delení. Práve nezávislá kombinácia chromozómov v meióze je základom tretieho zákona G. Mendela. Vzhľad zeleného hladkého a žltého vráskavého semena hrachu v druhej generácii krížením rastlín so žltými hladkými a zelenými vráskavými semenami je príkladom kombinačnej variability.
Vzájomná výmena oblastí homológnych chromozómov alebo kríženie. Vytvára nové väzbové skupiny, to znamená, že slúži ako dôležitý zdroj genetickej rekombinácie alel. Keď sú rekombinantné chromozómy v zygote, prispievajú k vzniku znakov, ktoré sú pre každého z rodičov netypické.
Náhodná kombinácia gamét počas oplodnenia.
Tieto zdroje kombinačnej variability pôsobia nezávisle a súčasne, pričom zaisťujú neustále „miešanie“ génov, čo vedie k vzniku organizmov s odlišným genotypom a fenotypom (samotné gény sa nemenia). Nové kombinácie génov sa však pri prechode z generácie na generáciu celkom ľahko degradujú.
Príklad kombinačnej variability. Kvetina nočnej krásy má gén pre červené okvetné lístky A a gén pre biele a. Telo Aa má ružové okvetné lístky. Nočná kráska teda nemá ružový gén; ružová sa objaví, keď dôjde k kombinácii (kombinácie) červeného a bieleho génu.
Osoba má dedičnú chorobu kosáčikovitú anémiu. AA je norma, aa je smrť, Aa je SKA. Pri SKA človek nemôže tolerovať zvýšenú fyzickú námahu, pričom netrpí maláriou, to znamená, že pôvodca malárie Plasmodium malaria sa nemôže živiť nesprávnym hemoglobínom. Táto funkcia je užitočná v rovníkovom páse; neexistuje na to žiadny gén, vzniká kombináciou génov A a a.
Dedičná variácia je teda posilnená kombinačnou variáciou. Po vzniku sa jednotlivé mutácie ocitnú v blízkosti iných mutácií, sú súčasťou nových genotypov, to znamená, že vzniká mnoho kombinácií alel. Každý jedinec je geneticky jedinečný (s výnimkou identických dvojčiat a jedincov, ktoré vznikli v dôsledku nepohlavnej reprodukcie klonu s jedinou bunkou ako predkom). Ak teda predpokladáme, že v každom páre homológnych chromozómov je iba jeden pár alelických génov, potom pre osobu, ktorej haploidná sada chromozómov je 23, bude počet možných genotypov 3 až 23 stupňov. Taký obrovský počet genotypov je 20 -násobkom počtu všetkých ľudí na Zemi. V skutočnosti sa však homológne chromozómy líšia v niekoľkých génoch a výpočet nezohľadňuje jav kríženia. . Preto je počet možných genotypov vyjadrený v astronomických číslach a dá sa sebavedomo tvrdiť, že výskyt dvoch identických ľudí je prakticky nepravdepodobný (s výnimkou identických dvojčiat, ktoré vznikli z rovnakého oplodneného vajíčka). Preto predovšetkým vyplýva z možnosti spoľahlivej identifikácie osoby podľa zvyškov živých tkanív, potvrdenia alebo vylúčenia otcovstva.
Výmena génov v dôsledku kríženia chromozómov v prvom delení meiózy, nezávislej a náhodnej rekombinácie chromozómov pri meióze a šance na fúziu gamét v sexuálnom procese sú tri faktory, ktoré zaisťujú existenciu kombinačnej variability. Mutačná variabilita samotného genotypu.
Mutácie sú náhle dedičné zmeny v genetickom materiáli, ktoré vedú k zmene určitých charakteristík organizmu.
Hlavné ustanovenia mutačnej teórie vyvinul vedec G. De Vries v roku 1901 – 1903 a varte do tohto:
Mutácie sa objavujú náhle, skokovo, ako diskrétne zmeny vlastností;
Na rozdiel od nededičných zmien sú mutácie kvalitatívne zmeny, ktoré sa dedia z generácie na generáciu;
Mutácie sa prejavujú rôznymi spôsobmi a môžu byť prospešné aj škodlivé, dominantné aj recesívne;
Pravdepodobnosť detekcie mutácií závisí od počtu vyšetrených jedincov;
Podobné mutácie sa môžu opakovať;
Mutácie sú nesmerové (spontánne), to znamená, že akákoľvek časť chromozómu môže mutovať, čo spôsobuje zmeny v menších aj vitálnych funkciách.
Takmer každá zmena štruktúry alebo počtu chromozómov, v ktorých si bunka zachováva schopnosť reprodukovať sa, spôsobuje dedičnú zmenu vlastností organizmu.
Podľa povahy zmeny genómu, to znamená, že sa rozlišuje súbor génov obsiahnutých v haploidnom súbore chromozómov, génové, chromozomálne a genómové mutácie.
Génové alebo bodové mutácie sú výsledkom zmeny nukleotidovej sekvencie v molekule DNA v rámci jedného génu.
Takáto zmena v géne sa reprodukuje počas transkripcie v štruktúre i-RNA; vedie k zmene v sekvencii aminokyselín v polypeptidovom reťazci vytvorenom počas translácie na ribozómoch. V dôsledku toho sa syntetizuje ďalší proteín, čo vedie k zmene zodpovedajúcej charakteristiky organizmu. Toto je najbežnejší typ mutácie a najdôležitejší zdroj dedičných variácií v organizmoch.
Chromozomálne mutácie (prestavby alebo aberácie) sú zmeny v štruktúre chromozómov, ktoré je možné identifikovať a študovať pod svetelným mikroskopom.
Sú známe preusporiadania rôznych typov:
nedostatok – strata koncov chromozómu;
Vymazanie – strata časti chromozómu v jeho strednej časti;
Duplikácia – dvojité alebo viacnásobné opakovanie génov lokalizovaných v určitej časti chromozómu;
Inverzia – rotácia chromozómovej časti o 180 °, v dôsledku čoho sú gény v tejto sekcii umiestnené v sekvencii opačnej k obvyklej;
Translokácia – zmena polohy ktorejkoľvek časti chromozómu v chromozómovej sade. Najbežnejší typ translokácie je recipročný, v ktorom dochádza k výmene oblastí medzi dvoma nehomológnymi chromozómami. Časť chromozómu môže zmeniť svoju polohu bez vzájomnej výmeny, zostať v tom istom chromozóme alebo byť zahrnutá v inom.
Genomické mutácie - zmena počtu chromozómov v genóme buniek v organizme. Tento jav sa vyskytuje v dvoch smeroch: k zvýšeniu počtu celých haploidných súborov (polyploidia) a k strate alebo začleneniu jednotlivých chromozómov (aneuploidia).
Polyploidia – viacnásobné zvýšenie haploidnej sady chromozómov. Bunky s rôznym počtom haploidných súborov chromozómov sa nazývajú triploidné (3 n), tetraploidové (4 n), hexaploidné (6 n), oktaploidové (8 n) atď. Polyploidy sa najčastejšie tvoria, keď je poradie divergencie chromozómov na bunkové póly počas meiózy sú narušené alebo mitóza. Polyploidia vedie k zmene charakteristík organizmu, a preto je dôležitým zdrojom variability v evolúcii a selekcii, najmä v rastlinách. Je to spôsobené tým, že hermafroditizmus (samoopelenie), apomixis (partenogenéza) a vegetatívna reprodukcia sú v rastlinných organizmoch veľmi rozšírené. Preto je asi tretina druhov rastlín bežných na našej planéte, – polyploidy a v ostro kontinentálnych podmienkach vysokohorských Pamírov rastie až 85% polyploidov. Takmer všetky kultivované rastliny sú tiež polyploidy, ktoré na rozdiel od svojich divokých príbuzných majú väčšie kvety, plody a semená a v zásobných orgánoch (stonka, hľuzy) sa hromadí viac živín. Polyploidi sa ľahšie prispôsobujú nepriaznivým životným podmienkam, ľahšie tolerujú nízke teploty a sucho. Preto sú rozšírené v severných a vysokých horských oblastiach.