Dešifriranje genoma čimpanze i usporedba s ljudskim genomom. Drosophila s drevnim genom opovrgla je jednu od klasičnih teorija evolucije
Uz 50. obljetnicu otkrića strukture DNK
A.V. Zelenin
BILJNI GENOM
A. V. Zelenin
Zelenin Aleksandar Vladimirovič- doktor bioloških znanosti,
Voditelj laboratorija, Institut za molekularnu biologiju nazvan. V.A. Engelhardt RAS.
Impresivna postignuća programa Ljudski genom, kao i uspjeh rada na dešifriranju takozvanih ultra-malih (virusi), malih (bakterije, kvasci) i srednjih (valjkasti crvi, drozofile) genoma, omogućili su prijeći na opsežnu studiju velikih i ekstra velikih biljnih genoma. Na skupu o biljnoj genomici održanom 1997. godine u SAD-u [,] naglašena je hitna potreba za detaljnim proučavanjem genoma ekonomski najvažnijih biljaka. Tijekom godina od tada postignuti su nedvojbeni uspjesi na ovom području. Godine 2000. pojavila se publikacija o potpunom sekvencioniranju (uspostavljanje linearnog slijeda nukleotida sve nuklearne DNA) genoma male gorušice - Arabidopsis, a 2001. - o preliminarnom (nacrtu) sekvencioniranja genoma riže. Rad na sekvencioniranju genoma velikih i ultravelikih biljaka (kukuruz, raž, pšenica) više puta je izvješćivan, ali te poruke nisu sadržavale konkretne informacije i bile su prije izjave namjere.
Očekuje se da će dešifriranje biljnih genoma otvoriti široke perspektive znanosti i praksi. Prije svega, identifikacija novih gena i lanac njihove genetske regulacije značajno će povećati produktivnost biljaka primjenom biotehnoloških pristupa. Otkriće, izolacija, reprodukcija (kloniranje) i sekvenciranje gena odgovornih za tako važne funkcije biljnog organizma kao što su reprodukcija i produktivnost, procesi varijabilnosti, otpornost na nepovoljne čimbenike okoliša, kao i homologno sparivanje kromosoma, povezano je s pojavom novih mogućnosti za poboljšanje selekcijskog procesa. Konačno, izolirani i klonirani geni mogu se koristiti za dobivanje transgenih biljaka s temeljno novim svojstvima i analizu mehanizama regulacije aktivnosti gena.
O važnosti proučavanja biljnih genoma govori i činjenica da je do sada broj lokaliziranih, kloniranih i sekvenciranih biljnih gena malen i prema različitim procjenama varira između 800 i 1200. To je 10-15 puta manje nego npr. na primjer, kod ljudi.
Sjedinjene Američke Države ostaju nedvojbeni lider u velikom istraživanju genoma biljaka, iako se intenzivna istraživanja genoma riže provode u Japanu, a posljednjih godina iu Kini. Osim američkih laboratorija, europske istraživačke skupine aktivno su sudjelovale u dešifriranju genoma Arabidopsis. Očigledno vodstvo Sjedinjenih Američkih Država izaziva ozbiljnu zabrinutost europskih znanstvenika, što su oni jasno iskazali na skupu znakovito nazvanom “Izgledi za genomiku u postgenomskoj eri” održanom u Francuskoj krajem 2000. godine. Napredak američke znanosti u proučavanju genoma poljoprivrednih biljaka i stvaranju transgenih biljnih oblika, prema europskim znanstvenicima, prijeti da će u ne tako dalekoj budućnosti (od dva do pet desetljeća), kada će porast populacije dovesti čovječanstvo u lice s opće prehrambene krize, europsko gospodarstvo i znanost postat će ovisni o američkoj tehnologiji. S tim u vezi, najavljeno je stvaranje francusko-njemačkog znanstvenog programa za proučavanje biljnih genoma (Plantgene) i ulaganje značajnih sredstava u njega.
Očito, problemi genomike biljaka trebali bi privući veliku pozornost ruskih znanstvenika i organizatora znanosti, kao i upravnih tijela, budući da govorimo ne samo o znanstvenom prestižu, već io nacionalnoj sigurnosti zemlje. Za jedno ili dva desetljeća hrana će postati najvažniji strateški resurs.
TEŠKOĆE U PROUČAVANJU BILJNIH GENOMA
Proučavanje genoma biljaka mnogo je složeniji zadatak od proučavanja genoma ljudi i drugih životinja. To je zbog sljedećih okolnosti:
ogromne veličine genoma, koji dosežu desetke pa čak i stotine milijardi parova nukleotida (bp) za pojedine biljne vrste: genomi glavnih gospodarski važnih biljaka (osim riže, lana i pamuka) su ili blizu veličine ljudskog genoma ili ga premašuju mnogo puta (stol);STUDIJE KROMOSOMSKOG GENOMAOštre fluktuacije u broju kromosoma u različitim biljkama - od dva u nekim vrstama do nekoliko stotina u drugima, a nije moguće utvrditi strogu korelaciju između veličine genoma i broja kromosoma;
Obilje poliploida (koji sadrže više od dva genoma po stanici) formira se sa sličnim ali ne identičnim genomima (alopoliploidija);
Ekstremno obogaćivanje biljnih genoma (do 99%) s "beznačajnom" (nekodirajućom, tj. koja ne sadrži gene) DNA, što uvelike komplicira spajanje (raspored u ispravnom redoslijedu) sekvenciranih fragmenata u zajednički veliki- dimenzionirana regija DNA (contig);
Nepotpuno (u usporedbi s genomima Drosophile, čovjeka i miša) morfološko, genetsko i fizičko mapiranje kromosoma;
Praktična nemogućnost izolacije pojedinačnih kromosoma u čistom obliku korištenjem metoda koje se obično koriste u tu svrhu za ljudske i životinjske kromosome (protočno sortiranje i uporaba staničnih hibrida);
Poteškoće kromosomskog mapiranja (određivanje položaja na kromosomu) pojedinačnih gena pomoću hibridizacije in situ, zbog visokog sadržaja "beznačajne" DNA u biljnim genomima i osobitosti strukturne organizacije biljnih kromosoma;
Evolucijska udaljenost biljaka od životinja, koja ozbiljno komplicira korištenje informacija dobivenih sekvenciranjem genoma ljudi i drugih životinja za proučavanje biljnih genoma;
Dug proces reprodukcije većine biljaka, što značajno usporava njihovu genetsku analizu.
Kromosomska (citogenetička) istraživanja genoma općenito, a posebno biljaka, imaju dugu povijest. Pojam "genom" predložen je za označavanje haploidnog (pojedinačnog) skupa kromosoma s genima koje sadrže u prvoj četvrtini 20. stoljeća, dakle mnogo prije nego što je utvrđena uloga DNK kao nositelja genetske informacije.
Opis genoma novog, prethodno neistraženog genetski višestaničnog organizma obično počinje proučavanjem i opisom kompletnog skupa njegovih kromosoma (kariotipa). To se, naravno, odnosi i na biljke od kojih veliki broj nije niti počeo proučavati.
Već u osvit kromosomskih studija genomi srodnih biljnih vrsta uspoređivani su na temelju analize mejotičke konjugacije (ujedinjenja homolognih kromosoma) kod međuvrsnih hibrida. Tijekom proteklih 100 godina, mogućnosti kromosomske analize dramatično su se proširile. Danas se za karakterizaciju biljnih genoma koriste naprednije tehnologije: razne varijante tzv. diferencijalnog bojenja, koje omogućuje identifikaciju pojedinačnih kromosoma na temelju morfoloških karakteristika; hibridizacija in situ, omogućavanje lokalizacije specifičnih gena na kromosomima; biokemijska istraživanja staničnih proteina (elektroforeza i imunokemija) i, konačno, skup metoda temeljenih na analizi kromosomske DNA do njezinog sekvenciranja.

Riža. 1. Kariotipovi žitarica: a - raž (14 kromosoma), b - tvrda pšenica (28 kromosoma), c - meka pšenica (42 kromosoma), d - ječam (14 kromosoma)Kariotipovi žitarica, prvenstveno pšenice i raži, proučavaju se dugi niz godina. Zanimljivo je da je kod različitih vrsta ovih biljaka broj kromosoma različit, ali uvijek višestruki od sedam. Pojedine vrste žitarica mogu se pouzdano identificirati prema njihovom kariotipu. Na primjer, genom raži sastoji se od sedam parova velikih kromosoma s intenzivno obojenim heterokromatskim blokovima na njihovim krajevima, koji se često nazivaju segmenti ili trake (Slika 1a). Genomi pšenice već imaju 14 i 21 par kromosoma (slika 1, b, c), a raspodjela heterokromatskih blokova u njima nije ista kao u kromosomima raži. Međusobno se razlikuju i pojedinačni genomi pšenice, označeni A, B i D. Povećanje broja kromosoma s 14 na 21 dovodi do oštre promjene svojstava pšenice, što se odražava u njihovim nazivima: durum, durum, ili makaroni, pšenični i meki, ili kruh, pšenični . Gen D, koji sadrži gene za proteine glutena, odgovoran je za stjecanje visokih pekarskih svojstava meke pšenice, što tijestu daje tzv. Upravo tom genomu pridaje se posebna pozornost u selekcijskom poboljšanju krušne pšenice. Još jedna žitarica s 14 kromosoma, ječam (slika 1, d), obično se ne koristi za izradu kruha, ali služi kao glavna sirovina za proizvodnju uobičajenih proizvoda kao što su pivo i viski.
Intenzivno se proučavaju kromosomi nekih samoniklih biljaka koje se koriste za poboljšanje kakvoće najvažnijih poljoprivrednih vrsta, primjerice divlji srodnici pšenice - Aegilops. Križanjem (slika 2) i selekcijom nastaju novi biljni oblici. Posljednjih godina, značajna poboljšanja u istraživačkim metodama omogućila su početak proučavanja genoma biljaka čije su značajke kariotipa (uglavnom male veličine kromosoma) učinile ranije nedostupnima za kromosomsku analizu. Tako su tek nedavno prvi put identificirani svi kromosomi pamuka, kamilice i lana.

Riža. 2. Kariotipovi pšenice i pšenično-Aegilops hibrida
a - heksaploidna obična pšenica ( Triticum astivum), koji se sastoji od A, B i O genoma; b - tetraploidna pšenica ( Triticum timopheevi), koji se sastoji od A i G genoma. sadrži gene otpornosti na većinu bolesti pšenice; c - hibridi Triticum astivum x Triticum timopheevi, otporan na pepelnicu i hrđu, jasno je vidljiva zamjena dijela kromosomaPRIMARNA STRUKTURA DNK
Kako se molekularna genetika razvijala, širio se i sam koncept genoma. Sada se ovaj pojam tumači iu klasičnom kromosomskom iu modernom molekularnom smislu: cjelokupni genetski materijal pojedinog virusa, stanice i organizma. Naravno, nakon proučavanja kompletne primarne strukture genoma (kako se često naziva kompletan linearni slijed baza nukleinskih kiselina) niza mikroorganizama i ljudi, postavilo se pitanje sekvenciranja genoma biljaka.
Od brojnih biljnih organizama za proučavanje su odabrana dva - Arabidopsis, predstavnik klase dikotiledona (veličina genoma 125 milijuna bp), i riža iz klase jednosupnica (420-470 milijuna bp). Ti su genomi mali u usporedbi s drugim biljnim genomima i sadrže relativno malo ponovljenih dijelova DNA. Takve značajke davale su nadu da će odabrani genomi biti dostupni za relativno brzo određivanje njihove primarne strukture.

Riža. 3. Arabidopsis - mala gorušica - mala biljka iz porodice krstašica ( Brassicaceae). Na prostoru jednakom površini jedne stranice našeg časopisa može se uzgajati do tisuću pojedinačnih organizama ArabidopsisOsnova za odabir Arabidopsisa nije bila samo mala veličina njegovog genoma, već i mala veličina organizma, što olakšava uzgoj u laboratorijskim uvjetima (slika 3). Uzeli smo u obzir njegov kratki reprodukcijski ciklus, zahvaljujući kojem je moguće brzo provesti pokuse križanja i selekcije, detaljnu genetiku, lakoću manipulacije promjenjivim uvjetima uzgoja (promjena sastava soli u tlu, dodavanje različitih hranjiva itd.) i ispitivanje djelovanja na biljke različitih mutagenih čimbenika i patogena (virusi, bakterije, gljivice). Arabidopsis nema ekonomsku vrijednost, stoga je njegov genom, zajedno s genomom miša, nazvan referentnim genomom ili, manje točno, modelnim genomom.*
* Pojava pojma "model genoma" u ruskoj literaturi rezultat je netočnog prijevoda engleske fraze model genome. Riječ "model" ne označava samo pridjev "model", već i imenicu "uzorak", "standard", "model". Ispravnije bi bilo govoriti o genomu uzorka, odnosno o referentnom genomu.Intenzivan rad na sekvencioniranju genoma Arabidopsis započeo je 1996. godine međunarodni konzorcij koji je uključivao znanstvene institucije i istraživačke skupine iz SAD-a, Japana, Belgije, Italije, Velike Britanije i Njemačke. U prosincu 2000. postale su dostupne opsežne informacije koje sažimaju određivanje primarne strukture genoma Arabidopsis. Za sekvenciranje smo koristili klasičnu, odnosno hijerarhijsku tehnologiju: prvo su proučavani pojedinačni mali dijelovi genoma, od kojih su napravljeni veći dijelovi (kontigi), au završnoj fazi struktura pojedinih kromosoma. Jezgra DNA genoma Arabidopsis raspoređena je među pet kromosoma. Godine 1999. objavljeni su rezultati sekvenciranja dvaju kromosoma, a objavom podataka o primarnoj strukturi preostala tri dovršeno je sekvenciranje cijelog genoma.
Od 125 milijuna parova nukleotida utvrđena je primarna struktura njih 119 milijuna, što je 92% cjelokupnog genoma. Pokazalo se da je samo 8% genoma Arabidopsis, koji sadrži velike blokove ponavljajućih dijelova DNK, nedostupno za proučavanje. U pogledu potpunosti i temeljitosti sekvenciranja eukariotskih genoma, Arabidopsis ostaje među prva tri prvaka zajedno s jednostaničnim organizmom kvasca Saccharomyces cerevisiae i višestanični životinjski organizam Caenorhabditis elegancija(vidi tablicu).
Oko 15 tisuća pojedinačnih gena koji kodiraju proteine pronađeno je u genomu Arabidopsis. Otprilike 12 tisuća njih sadržano je u dvije kopije po haploidnom (jednom) genomu, tako da je ukupan broj gena 27 tisuća.Broj gena u Arabidopsisu ne razlikuje se puno od broja gena u organizmima kao što su ljudi i miševi, ali veličina njegovog genoma 25-30 puta manja. Ova je okolnost povezana s važnim značajkama u strukturi pojedinačnih gena Arabidopsis i cjelokupne strukture njegovog genoma.
Geni Arabidopsis su kompaktni, sadržavaju samo nekoliko egzona (protein-kodirajućih regija), odvojenih kratkim (oko 250 bp) nekodirajućim dionicama DNA (intronima). Razmaci između pojedinih gena u prosjeku iznose 4,6 tisuća parova nukleotida. Usporedbe radi, ističemo da ljudski geni sadrže mnogo desetaka pa čak i stotina egzona i introna, a međugenske regije imaju veličine od 10 tisuća parova nukleotida ili više. Vjeruje se da je prisutnost malog kompaktnog genoma pridonijela evolucijskoj stabilnosti Arabidopsis, budući da je njegova DNA postala manje meta za razne štetne agense, posebno za uvođenje ponavljajućih fragmenata DNA nalik virusu (transpozona) u genom.
Druge molekularne značajke genoma Arabidopsis uključuju obogaćivanje egzona gvaninom i citozinom (44% u egzonima i 32% u intronima) u usporedbi sa životinjskim genima, kao i prisutnost dva puta ponovljenih (dupliciranih) gena. Vjeruje se da se to udvostručenje dogodilo kao rezultat četiri istodobna događaja, koji su se sastojali u udvostručavanju (ponavljanju) dijela gena Arabidopsis, odnosno spajanju povezanih genoma. Ovi događaji, koji su se dogodili prije 100-200 milijuna godina, manifestacija su opće tendencije poliploidizacije (višestruko povećanje broja genoma u organizmu), karakteristične za genome biljaka. Međutim, neke činjenice pokazuju da u Arabidopsis duplicirani geni nisu identični i funkcioniraju drugačije, što može biti posljedica mutacija u njihovim regulatornim regijama.
Još jedan objekt potpunog sekvenciranja DNK bila je riža. Genom ove biljke također je mali (12 kromosoma, što ukupno daje 420-470 milijuna bp), samo 3,5 puta veći od genoma Arabidopsis. Međutim, za razliku od Arabidopsis, riža ima golemu gospodarsku važnost, jer je temelj prehrane za više od polovice čovječanstva, stoga nisu samo milijarde potrošača vitalno zainteresirane za poboljšanje njezinih svojstava, već i višemilijunska vojska ljudi koji su aktivno uključeni u vrlo radno intenzivan proces njegova uzgoja.
Neki istraživači počeli su proučavati genom riže još 80-ih godina prošlog stoljeća, ali taj je rad dosegao ozbiljne razmjere tek 90-ih. Godine 1991. u Japanu je stvoren program za dešifriranje strukture genoma riže, kombinirajući napore mnogih istraživačkih skupina. Godine 1997. na temelju tog programa organiziran je Međunarodni projekt genoma riže. Njegovi sudionici odlučili su usredotočiti svoje napore na sekvenciranje jedne od podvrsta riže ( Oriza sativajaponica), u čijem proučavanju je do tada već bio postignut značajan napredak. Program Ljudski genom postao je ozbiljan poticaj i, slikovito rečeno, zvijezda vodilja za takav rad.
U sklopu ovog programa testirana je strategija “kromosomske” hijerarhijske podjele genoma kojom su sudionici međunarodnog konzorcija dešifrirali genom riže. Međutim, ako su pri proučavanju ljudskog genoma frakcije pojedinih kromosoma izolirane različitim tehnikama, onda je materijal specifičan za pojedine kromosome riže i njihove pojedinačne dijelove dobiven laserskom mikrodisekcijom (izrezivanjem mikroskopskih objekata). Na stakalcu mikroskopa na kojem se nalaze kromosomi riže, pod utjecajem laserske zrake izgara sve osim kromosoma ili njegovih dijelova namijenjenih analizi. Preostali materijal koristi se za kloniranje i sekvenciranje.
Objavljena su brojna izvješća o rezultatima sekvencioniranja pojedinih fragmenata genoma riže, provedenog s visokom točnošću i detaljima karakterističnim za hijerarhijsku tehnologiju. Vjerovalo se da će određivanje potpune primarne strukture genoma riže biti dovršeno do kraja 2003. - sredine 2004. godine i da će rezultati, zajedno s podacima o primarnoj strukturi genoma Arabidopsis, biti široko korišteni u komparativnoj genomici. drugih biljaka.
Međutim, početkom 2002. dvije istraživačke skupine - jedna iz Kine, druga iz Švicarske i Sjedinjenih Država - objavile su rezultate potpunog grubog (grubog) sekvenciranja genoma riže, izvedenog korištenjem tehnologije totalnog kloniranja. Za razliku od postupne (hijerarhijske) studije, ukupni pristup temelji se na istovremenom kloniranju cijele genomske DNA u jednom od virusnih ili bakterijskih vektora i dobivanju značajnog (ogromnog za srednje i velike genome) broja pojedinačni klonovi koji sadrže različite segmente DNA. Na temelju analize ovih sekvenciranih odsječaka i preklapanja identičnih krajnjih odsječaka DNK formira se kontig – lanac spojenih DNK sekvenci. Opći (ukupni) kontig predstavlja primarnu strukturu cijelog genoma ili barem pojedinog kromosoma.
U takvom shematskom prikazu strategija potpunog kloniranja izgleda nekomplicirana. Zapravo, nailazi na ozbiljne poteškoće povezane s potrebom dobivanja ogromnog broja klonova (opće je prihvaćeno da se genom ili njegova regija koja se proučava mora preklapati klonovima najmanje 10 puta), gigantskim volumenom sekvenciranja i iznimno složen posao spajanja klonova koji zahtijeva sudjelovanje bioinformatičara. Ozbiljna prepreka potpunom kloniranju je raznolikost ponavljajućih regija DNK, čiji se broj, kao što je već spomenuto, naglo povećava s povećanjem veličine genoma. Stoga se strategija totalnog sekvenciranja prvenstveno koristi u proučavanju genoma virusa i mikroorganizama, iako je uspješno primijenjena u proučavanju genoma višestaničnog organizma, Drosophile.
Rezultati totalnog sekvencioniranja ovog genoma "superiponirani" su na ogroman niz informacija o njegovoj kromosomskoj, genskoj i molekularnoj strukturi dobivenih tijekom gotovo 100-godišnjeg razdoblja proučavanja Drosophile. Pa ipak, u pogledu stupnja sekvenciranja, genom Drosophila (66% ukupne veličine genoma) znatno je inferioran genomu Arabidopsis (92%), unatoč prilično sličnim veličinama - 180 milijuna odnosno 125 milijuna parova nukleotida. . Stoga je nedavno predloženo da se tehnologija korištena za sekvenciranje genoma Drosophile nazove miješanom.
Za sekvenciranje genoma riže, gore spomenute istraživačke skupine uzele su dvije njezine podvrste, koje se najviše uzgajaju u azijskim zemljama - Oriza slina L. ssp indicaj I Oriza slina L. sspjaponica. Rezultati njihova istraživanja u mnogočemu se podudaraju, ali i razlikuju. Tako su predstavnici obje skupine izjavili da su postigli preklapanje kontiga od približno 92-93% genoma. Pokazalo se da je oko 42% genoma riže predstavljeno kratkim DNA ponavljanjima koja se sastoje od 20 parova nukleotida, a većina mobilnih DNA elemenata (transpozona) nalazi se u intergenskim regijama. Međutim, podaci o veličini genoma riže značajno variraju.
Za japansku podvrstu veličina genoma određena je na 466 milijuna parova nukleotida, a za indijsku podvrstu - 420 milijuna.Razlog ove razlike nije jasan. Ona može biti posljedica različitih metodoloških pristupa određivanju veličine nekodirajućeg dijela genoma, odnosno ne mora odražavati pravo stanje stvari. No moguće je da razlika od 15% u veličini proučavanih genoma doista postoji.
Druga ozbiljna razlika otkrivena je u broju otkrivenih gena: za japansku podvrstu - od 46 022 do 55 615 gena po genomu, a za indijsku podvrstu - od 32 000 do 50 000. Razlog za ovu razliku nije jasan.
Nepotpunost i nedosljednost dobivenih informacija uočena je u komentarima na objavljene članke. Također se nadamo da će praznine u poznavanju genoma riže biti eliminirane usporedbom podataka iz "grubog sekvenciranja" s rezultatima detaljnog, hijerarhijskog sekvenciranja koje su proveli sudionici u Međunarodnom projektu genoma riže.
USPOREDNA I FUNKCIONALNA GENOMIKA BILJA
Dobiveni opsežni podaci, od kojih je polovica (rezultati kineske skupine) javno dostupni, nedvojbeno otvaraju široke perspektive kako za proučavanje genoma riže, tako i za genomiku biljaka općenito. Usporedba svojstava genoma Arabidopsis i riže pokazala je da se većina gena (do 80%) identificiranih u genomu Arabidopsis također nalazi u genomu riže, međutim, za otprilike polovicu gena pronađenih u riži, analozi ( ortolozi) još nisu pronađeni u genomu Arabidopsis. Istodobno je u genomu riže identificirano 98% gena čija je primarna struktura utvrđena za druge žitarice.
Značajna (gotovo dvostruka) razlika u broju gena u riži i Arabidopsisu je zbunjujuća. Istodobno, podaci iz grubog prijepisa genoma riže, dobiveni totalnim sekvenciranjem, praktički se ne uspoređuju s opsežnim rezultatima proučavanja genoma riže metodom hijerarhijskog kloniranja i sekvenciranja, odnosno onim što je učinjeno. za genom Drosophila nije postignut. Stoga ostaje nejasno odražava li razlika u broju gena u Arabidopsisu i riži pravo stanje stvari ili se objašnjava razlikama u metodološkim pristupima.
Za razliku od genoma Arabidopsis, informacije o genima blizanaca u genomu riže nisu dane. Moguće je da je njihova relativna brojnost veća u riži nego u Arabidopsisu. Ovu mogućnost posredno podupiru podaci o prisutnosti poliploidnih oblika riže. Veća jasnoća po ovom pitanju može se očekivati nakon završetka Međunarodnog projekta genoma riže i dobivanja detaljne slike primarne strukture DNK ovog genoma. Ozbiljne razloge za takvu nadu daje činjenica da je nakon objavljivanja radova o grubom sekvenciranju genoma riže naglo porastao broj publikacija o strukturi ovog genoma, posebice su se pojavile informacije o detaljnom sekvenciranju njegovih kromosoma. 1 i 4.
Poznavanje, barem približno, broja gena u biljkama od temeljne je važnosti za komparativnu biljnu genomiku. U početku se vjerovalo da, budući da su sve cvjetnice vrlo bliske jedna drugoj u svojim fenotipskim karakteristikama, njihovi bi genomi također trebali biti bliski. A ako proučavamo genom Arabidopsis, dobit ćemo informacije o većini genoma drugih biljaka. Neizravnu potvrdu ove pretpostavke daju rezultati sekvenciranja genoma miša, koji je iznenađujuće blizak ljudskom genomu (oko 30 tisuća gena, od kojih se samo 1 tisuću pokazalo različitim).
Može se pretpostaviti da razlog razlika u genomima Arabidopsis i riže leži u njihovoj pripadnosti različitim klasama biljaka - dikotiledonima i monokotiledonima. Da bismo razjasnili ovo pitanje, iznimno je poželjno poznavati barem okvirnu primarnu strukturu neke druge jednosupnice. Najrealniji kandidat mogao bi biti kukuruz čiji je genom približno jednak ljudskom genomu, ali ipak značajno manji od genoma ostalih žitarica. Prehrambena vrijednost kukuruza dobro je poznata.
Ogroman materijal dobiven sekvenciranjem genoma Arabidopsis i riže postupno postaje osnova za opsežna istraživanja biljnih genoma korištenjem komparativnih genomskih metoda. Takva istraživanja imaju opći biološki značaj, jer omogućuju utvrđivanje glavnih principa organizacije biljnog genoma kao cjeline i njihovih pojedinačnih kromosoma, identificiranje zajedničkih značajki strukture gena i njihovih regulatornih područja te razmatranje odnos između funkcionalno aktivnog (genskog) dijela kromosoma i različitih intergenskih regija DNA koje ne kodiraju proteine. Komparativna genetika također postaje sve važnija za razvoj ljudske funkcionalne genomike. Za komparativne studije sekvencionirani su genomi riba napuhača i miševa.
Ne manje važno je proučavanje pojedinih gena odgovornih za sintezu pojedinih proteina koji određuju specifične funkcije tijela. Upravo u detekciji, izolaciji, sekvencioniranju i utvrđivanju funkcije pojedinih gena leži praktični, prvenstveno medicinski, značaj programa Ljudski genom. Ovu je okolnost uočio prije nekoliko godina J. Watson, ističući da će program Ljudski genom biti dovršen tek kada se utvrde funkcije svih ljudskih gena.
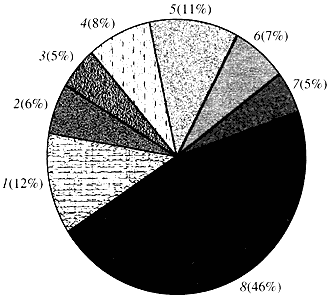
Riža. 4. Klasifikacija prema funkciji gena Arabidopsis
1 - geni za rast, diobu i sintezu DNA; 2 - geni sinteze RNA (transkripcija); 3 - geni za sintezu i modifikaciju proteina; 4 - geni za razvoj, starenje i smrt stanice; 5 - geni staničnog metabolizma i metabolizma energije; 6 - geni za međustaničnu interakciju i prijenos signala; 7 - geni za potporu drugim staničnim procesima; 8 - geni s nepoznatom funkcijomKada je riječ o funkciji biljnih gena, znamo manje od jedne desetine onoga što znamo o ljudskim genima. Čak i kod Arabidopsisa, čiji je genom mnogo više proučavan od ljudskog genoma, funkcija gotovo polovice njegovih gena ostaje nepoznata (slika 4). U međuvremenu, biljke, osim gena zajedničkih životinjama, imaju značajan broj gena specifičnih samo (ili barem pretežno) za njih. Riječ je o genima koji su uključeni u transport vode i sintezu staničnih stijenki, kojih nema u životinja, o genima koji osiguravaju stvaranje i funkcioniranje kloroplasta, fotosintezu, fiksaciju dušika i sintezu brojnih aromatskih proizvoda. Ovaj popis se može nastaviti, ali već je jasno koliko je težak zadatak funkcionalna genomika biljaka.
Kompletno sekvenciranje genoma daje približno istinitu informaciju o ukupnom broju gena određenog organizma, omogućuje više ili manje detaljne i pouzdane podatke o njihovoj strukturi da se smjeste u banke podataka te olakšava rad na izdvajanju i proučavanju pojedinačnih gena. Međutim, sekvenciranje genoma ne znači utvrđivanje funkcije svih gena.
Jedan od najperspektivnijih pristupa funkcionalne genomike temelji se na identificiranju radnih gena na kojima se događa transkripcija (čitanje) mRNA. Ovaj pristup, uključujući korištenje moderne tehnologije mikronizova, omogućuje istodobnu identifikaciju do desetaka tisuća funkcionalnih gena. Nedavno je ovim pristupom započelo proučavanje biljnih genoma. Za Arabidopsis je bilo moguće dobiti oko 26 tisuća pojedinačnih transkripata, što uvelike olakšava mogućnost utvrđivanja funkcije gotovo svih njegovih gena. U krumpiru je bilo moguće identificirati oko 20.000 tisuća aktivnih gena koji su važni za razumijevanje kako procesa rasta i formiranja gomolja, tako i procesa bolesti krumpira. Očekuje se da će ovo saznanje poboljšati otpornost jednog od najvažnijih prehrambenih proizvoda na patogene.
Logičan razvoj funkcionalne genomike je proteomika. Ovo novo područje znanosti proučava proteome, koji se obično odnose na kompletan skup proteina u stanici u određenom trenutku. Ovaj skup proteina, koji odražava funkcionalno stanje genoma, mijenja se cijelo vrijeme, dok genom ostaje nepromijenjen.
Proučavanje proteina dugo se koristilo za donošenje prosudbi o aktivnosti biljnih genoma. Kao što je poznato, enzimi koji se nalaze u svim biljkama razlikuju se u redoslijedu aminokiselina u pojedinim vrstama i sortama. Takvi enzimi, s istom funkcijom, ali različitim slijedom pojedinih aminokiselina, nazivaju se izoenzimi. Imaju različita fizikalno-kemijska i imunološka svojstva (molekularna težina, naboj), koja se mogu detektirati kromatografijom ili elektroforezom. Ovim se metodama već dugi niz godina uspješno proučava tzv. genetski polimorfizam, odnosno razlike između organizama, sorti, populacija, vrsta, posebice pšenice i srodnih oblika žitarica. Međutim, u posljednje vrijeme, zbog brzog razvoja metoda analize DNA, uključujući i sekvencioniranje, proučavanje polimorfizma proteina zamijenjeno je proučavanjem polimorfizma DNA. Međutim, izravno proučavanje spektra skladišnih proteina (prolamini, glijadini i dr.), koji određuju osnovna nutritivna svojstva žitarica, ostaje važna i pouzdana metoda za genetičku analizu, selekciju i proizvodnju sjemena poljoprivrednih biljaka.
Poznavanje gena, mehanizama njihove ekspresije i regulacije iznimno je važno za razvoj biotehnologije i proizvodnju transgenih biljaka. Poznato je da impresivni uspjesi u ovom području izazivaju različite reakcije ekološke i medicinske zajednice. Međutim, postoji područje biljne biotehnologije u kojem se ti strahovi, ako ne potpuno neutemeljeni, onda, u svakom slučaju, čine beznačajnima. Riječ je o stvaranju transgenih industrijskih biljaka koje se ne koriste kao prehrambeni proizvodi. Indija je nedavno požnjela svoj prvi usjev transgenog pamuka koji je otporan na brojne bolesti. Postoje informacije o uvođenju posebnih gena koji kodiraju pigmentne proteine u genom pamuka i proizvodnji pamučnih vlakana koja ne zahtijevaju umjetno bojanje. Još jedna industrijska kultura koja može biti predmet učinkovitog genetskog inženjeringa je lan. Nedavno se raspravljalo o njegovoj upotrebi kao alternative pamuku za tekstilne sirovine. Ovaj problem je izuzetno važan za našu zemlju, koja je izgubila vlastite izvore pamučne sirovine.
IZGLEDI ZA PROUČAVANJE BILJNIH GENOMA
Očito je da će se strukturna istraživanja biljnih genoma temeljiti na pristupima i metodama komparativne genomike koristeći rezultate dešifriranja genoma Arabidopsis i riže kao glavni materijal. Značajnu ulogu u razvoju komparativne genomike biljaka bez sumnje će odigrati informacije koje će prije ili kasnije dati ukupno (grubo) sekvenciranje genoma drugih biljaka. U ovom slučaju komparativna genomika biljaka temeljit će se na utvrđivanju genetskih odnosa između pojedinih lokusa i kromosoma koji pripadaju različitim genomima. Nećemo govoriti toliko o općoj genomici biljaka, koliko o selektivnoj genomici pojedinih kromosomskih lokusa. Tako je nedavno pokazano da se gen odgovoran za vernalizaciju nalazi u lokusu VRn-AI kromosoma 5A heksaploidne pšenice i lokusu Hd-6 kromosoma 3 riže.
Razvoj ovih studija bit će snažan poticaj za identifikaciju, izolaciju i sekvenciranje mnogih funkcionalno važnih biljnih gena, posebno gena odgovornih za otpornost na bolesti, otpornost na sušu i prilagodljivost različitim uvjetima uzgoja. Sve više će se koristiti funkcionalna genomika, koja se temelji na masovnoj identifikaciji (screeningu) gena koji funkcioniraju u biljkama.
Možemo predvidjeti daljnja poboljšanja kromosomskih tehnologija, prvenstveno metode mikrodisekcije. Njegova uporaba dramatično proširuje mogućnosti genomskog istraživanja bez potrebe za velikim troškovima, kao što je sekvencioniranje ukupnog genoma. Metoda lokalizacije pojedinih gena na biljnim kromosomima pomoću hibridizacije postat će sve raširenija. in situ. Trenutačno je njegova uporaba ograničena ogromnim brojem ponavljajućih sekvenci u biljnom genomu, a vjerojatno i osobitostima strukturne organizacije biljnih kromosoma.
U doglednoj budućnosti kromosomske tehnologije će također postati od velike važnosti za evolucijsku genomiku biljaka. Te tehnologije, koje su relativno jeftine, omogućuju brzu procjenu intra- i interspecifične varijabilnosti i proučavanje složenih alopoliploidnih genoma tetraploidne i heksaploidne pšenice i tritikalea; analizirati evolucijske procese na kromosomskoj razini; istražiti nastanak sintetskih genoma i uvođenje (introgresiju) stranog genetskog materijala; identificirati genetske odnose između pojedinih kromosoma različitih vrsta.
Za karakterizaciju genoma koristit će se proučavanje kariotipa biljaka klasičnim citogenetskim metodama, obogaćenim molekularno-biološkom analizom i računalnim tehnologijama. To je posebno važno za proučavanje stabilnosti i varijabilnosti kariotipa na razini ne samo pojedinačnih organizama, već i populacija, sorti i vrsta. Naposljetku, teško je zamisliti kako se može procijeniti broj i spektar kromosomskih preraspodjela (aberacija, mostova) bez korištenja metoda diferencijalnog bojenja. Takva su istraživanja iznimno obećavajuća za praćenje okoliša na temelju stanja biljnog genoma.
U modernoj Rusiji malo je vjerojatno da će se provesti izravno sekvencioniranje biljnih genoma. Takav rad, koji zahtijeva velika ulaganja, neodrživ je za naše sadašnje gospodarstvo. U međuvremenu, informacije o strukturi genoma Arabidopsis i riže, dobivene svjetskom znanošću i dostupne u međunarodnim bankama podataka, dovoljne su za razvoj domaće biljne genomike. Moguće je predvidjeti širenje istraživanja biljnih genoma temeljenih na pristupima komparativne genomike za rješavanje specifičnih problema uzgoja i proizvodnje usjeva, kao i za proučavanje podrijetla različitih biljnih vrsta od gospodarskog značaja.
Može se pretpostaviti da će u domaćoj oplemenjivačkoj praksi i biljogojstvu biti široko korišteni genomski pristupi poput genetske tipizacije (RELF, RAPD, AFLP analize i dr.), koji su prilično pristupačni za naš budžet. Paralelno s izravnim metodama određivanja polimorfizma DNA, pristupi koji se temelje na proučavanju polimorfizma proteina, prvenstveno skladišnih proteina žitarica, koristit će se za rješavanje problema genetike i oplemenjivanja bilja. Kromosomske tehnologije bit će široko korištene. Relativno su jeftini, a njihov razvoj zahtijeva prilično umjerena ulaganja. U području istraživanja kromosoma domaća znanost nije niža od svjetske.
Treba naglasiti da je naša znanost dala značajan doprinos u formiranju i razvoju genomike biljaka [,].
Temeljnu ulogu odigrao je N.I. Vavilov (1887-1943).
U molekularnoj biologiji i biljnoj genomici očit je pionirski doprinos A.N. Belozerski (1905-1972).
U području istraživanja kromosoma potrebno je istaknuti rad izvanrednog genetičara S.G. Navashin (1857-1930), koji je prvi otkrio satelitske kromosome u biljkama i dokazao da je moguće razlikovati pojedine kromosome po karakteristikama njihove morfologije.
Još jedan klasik ruske znanosti G.A. Levitsky (1878-1942) detaljno je opisao kromosome raži, pšenice, ječma, graška i šećerne repe, uveo u znanost pojam "kariotip" i razvio doktrinu o njemu.
Suvremeni stručnjaci, oslanjajući se na dostignuća svjetske znanosti, mogu dati značajan doprinos daljnjem razvoju biljne genetike i genomike.
Autor izražava svoju srdačnu zahvalnost akademiku Yu.P. Altukhovu za kritičku raspravu o članku i dragocjene savjete.Rad tima na čelu s autorom članka poduprli su Ruska zaklada za temeljna istraživanja (grantovi br. 99-04-48832; 00-04-49036; 00-04-81086), Program predsjednika Ruska Federacija za potporu znanstvenim školama (grantovi br. 00-115 -97833 i NSh-1794.2003.4) i Program Ruske akademije znanosti "Molekularni genetski i kromosomski markeri u razvoju suvremenih metoda selekcije i sjemena proizvodnja."
KNJIŽEVNOST
1. Zelenin A.V., Badaeva E.D., Muravenko O.V. Uvod u genomiku biljaka // Molecular biology. 2001. T. 35. str. 339-348. 2. Olovka E. Bonanza za biljnu genomiku // Science. 1998. V. 282. P. 652-654. 3. Genomika biljaka // Proc. Natl. Akad. Sci. SAD. 1998. V. 95. P. 1962-2032. 4. Kartel N.A. i tako dalje. Genetika. Enciklopedijski rječnik. Minsk: Technologia, 1999. 5. Badaeva E.D., Friebe B., Gill B.S. 1996. Diferencijacija genoma kod Aegilopsa. 1. Distribucija visoko ponavljajućih sekvenci DNA na kromosomima diploidnih vrsta // Genome. 1996. V. 39. P. 293-306. Povijest analize kromosoma // Biol. membrane. 2001. T. 18. str. 164-172.
Za dva najautoritativnija svjetska znanstvena časopisa - britanski Nature i američki Science - izuzetna je rijetkost da značajan dio svojih sljedećih brojeva istovremeno posvete istoj temi. A ako se dogodi, to ukazuje na iznimnu važnost ove teme. Stoga je objavljivanje 12 članaka odjednom posvećenih dekodiranju genoma čimpanze i njegovoj usporedbi s ljudskim genomom, naravno, nesvakidašnji događaj.
Osnovan je međunarodni konzorcij za provedbu projekta mapiranja i komparativne analize genoma čimpanze. U njemu je sudjelovalo 67 znanstvenika iz 23 znanstvene institucije u 5 zemalja – SAD-u, Izraelu, Španjolskoj, Italiji i Njemačkoj. Rad su koordinirali genetičari sa Sveučilišta Harvard i Massachusetts Institute of Technology u Bostonu. A krv za DNK analizu osigurao je mladi mužjak čimpanze po imenu Clint, stanovnik jednog od ograđenih prostora u Nacionalnom centru za istraživanje primata Yerkes u Atlanti, Georgia. Nažalost, u siječnju ove godine donor je preminuo od akutnog zatajenja srca na vrhuncu života, u 24. godini života. Njegov je kostur sada izložen u muzeju Field u Chicagu. No, najvažnija vrijednost koju je čovječanstvo naslijedilo od Clinta je dio njegove krvi, koja je poslužila kao izvorni materijal za dešifriranje i analizu genoma čimpanze. Sada su se primati pridružili popisu organizama čiji je genetski materijal u potpunosti mapiran. Ovaj popis danas već uključuje stotine stavki: tu su i plijesni, bakterije, uključujući uzročnike opasnih zaraznih bolesti (antraks, tularemija, kuga, tifus), i biljke (riža, kavovo drvo), i insekti (malarični komarac), i ptice (npr. piletina) i sisavci (miš, štakor, pas, svinja, krava). Međutim, čovjekoliki majmuni zauzimaju, naravno, vrlo posebno mjesto na ovom popisu. Prema Robertu Waterstonu, direktoru genomskih istraživanja na Sveučilištu Washington Graduate School of Medicine u Seattleu, "proučavanje čimpanza, kao najbližih živih srodnika ljudima na Zemlji, može nam dati najviše informacija o nama samima." No, prije nego što prijeđem na raspravu o rezultatima koje su dobili znanstvenici, dopustit ću si malu digresiju - ili, ako želite, podsjetnik - kako bi bilo jasnije o čemu zapravo govorimo.
Kao što znate, bilo koji živi organizam sastoji se od stanica, au jezgri svake stanice nalazi se isti skup genetskih informacija karakterističnih za određenu biološku vrstu. Taj se skup naziva genom. Kromosomi su nositelji genetskih informacija. Kromosom je molekula deoksiribonukleinske kiseline (skraćeno DNK) i sastoji se od dva dugačka polinukleotidna lanca upletena jedan oko drugoga i međusobno povezana takozvanim vodikovim vezama. Ova se molekula naziva dvostruka spirala, a može se donekle pojednostavljeno zamisliti kao ljestve od upletenog užeta. Različite vrste životinja imaju različit broj kromosoma. Dakle, ljudski genom sastoji se od 23 para kromosoma – u svakom paru jedan kromosom dolazi od oca, drugi od majke. Vinska mušica - Drosophila - sadrži 4 para kromosoma u staničnoj jezgri, dok npr. bakterije imaju samo jedan nespareni kromosom. Geni se nalaze na kromosomima u strogo određenim područjima - svojevrsne jedinice nasljeđa. Kemijski, geni se sastoje od molekula 4 dušična spoja – adenina, citozina, gvanina i timina. Ove takozvane nukleotidne baze se ponavljaju u strogo definiranom redoslijedu, tvoreći parove adenin-timin i gvanin-citozin. Jedan gen može sadržavati od nekoliko tisuća do više od dva milijuna nukleotidnih baza. Njihov slijed određuje specifične funkcije svakog specifičnog gena.
Slikovito, genom se može zamisliti na sljedeći način: stanična jezgra je knjižnica u kojoj su pohranjene upute za osiguranje života; kromosomi igraju ulogu polica za knjige; na policama su knjige – molekule DNK; geni su poglavlja unutar knjiga, a nukleotidne baze - adenin, timin, gvanin i citozin, koje se obično označavaju početnim slovima njihovih imena A, T, G i C - upravo je to abeceda kojom je ispisan tekst genoma. napisano. Ljudski genom je, na primjer, lanac od 3 milijarde 200 milijuna slova.
Ali činjenica da geni postoje i da rade nije dovoljna: oni moraju djelovati na različite načine, osiguravajući određene specifične funkcije. Uostalom, stanice različitih organa i tkiva - recimo, kože, jetre, srca i mozga - nevjerojatno se razlikuju jedna od druge. U međuvremenu, jezgra svakog od njih sadrži isti skup gena. Sve je u aktivnosti gena: neki geni rade u nekim stanicama, a drugi u drugima. Dakle, kromosomi su nositelji ne samo gena, već i onih proteinskih čimbenika koji kontroliraju njihove funkcije. Taj skup gena, zajedno s regulatornim elementima, čini strukturu unutar stanice koja osigurava sve potrebne funkcije.
A sada, naoružani tim saznanjima, vratimo se rezultatima koji su dobiveni tijekom dešifriranja genoma čimpanze. Iz očitih razloga, najveći interes stručnjaka i šire javnosti predstavlja katalog onih razlika u genetskim kodovima čimpanza i ljudi koje su se akumulirale u proteklih 6 milijuna godina, od evolucijskih putova dviju vrsta koje su imale zajedničkog pretka odvojeni. Svante Pääbo, istraživač na Institutu Max Planck za evolucijsku antropologiju u Leipzigu i jedan od sudionika projekta, procjenjuje dobivenu bazu podataka na sljedeći način:
Riječ je o iznimno korisnom alatu koji će nam pomoći pronaći odgovor na pitanje koje genetske mutacije objašnjavaju upečatljivu razliku između čovjeka kao biološke vrste i svih ostalih životinjskih vrsta. Jedan od smjerova ove potrage svodi se na pokušaj utvrđivanja odnosa između genetskih razlika i aktivnosti pojedinih gena.
Prije svega, treba napomenuti da su dobiveni podaci iznenadili stručnjake. Glavno iznenađenje je da se pokazalo da je genom čimpanze 98,8 posto identičan ljudskom genomu. Grubo govoreći, genetska sličnost između ljudi i čimpanza je 10 puta veća nego između miševa i štakora. Amatere će vrlo vjerojatno iznenaditi tako velika sličnost, ta gotovo potpuna istovjetnost genoma, no znanstvenike je iznenadilo upravo suprotno: činjenica da se razlika pokazala prilično značajnom. Štoviše, ova brojka - slučajnost od 98,8 posto - ne odražava u potpunosti stanje stvari. Dobiva se usporedbom pojedinačnih slova genetskog koda u kodirajućoj DNK. Ovdje su znanstvenici izbrojali 35 milijuna odstupanja, što je iznosilo 1,2 posto cjelokupnog genoma čimpanze koji ima oko 3 milijarde i 100 milijuna parova nukleotida. Ali to nije sve: također su otkrivene značajne razlike u distribuciji onih sekvenci nukleotidnih baza koje tvore nekodirajuću, "sebičnu" DNK. Ova nepodudaranja činila su još 2,7 posto cjelokupnog genoma, što je ukupno gotovo 4 posto.
Ukupno su čimpanzama nedostajala 53 gena koja imaju ljudi. Konkretno, u genomu čimpanze nedostaju tri gena koji igraju ključnu ulogu u razvoju upale, za koju se zna da uzrokuje mnoge ljudske bolesti. S druge strane, čini se da su ljudi u procesu evolucije izgubili gen koji štiti životinje od Alzheimerove bolesti.
Najznačajnije razlike odnose se na gene koji reguliraju imunološki sustav. Prema profesoru Evanu Eichleru, suradniku na Medicinskom fakultetu Sveučilišta Washington u Seattleu, to ukazuje da su se čimpanze i ljudi morali suočiti s različitim patogenima i boriti protiv različitih bolesti tijekom svog evolucijskog razvoja. Svante Pääbo objašnjava:
Prije svega, zapitali smo se koji segmenti DNK mogu dati uvid u podrijetlo pojedinih bolesti. Znamo da se neke od genetskih struktura koje uzrokuju određenu bolest nalaze i kod čimpanza i kod ljudi. Očigledno su ove strukture obje vrste naslijedile od zajedničkog pretka. Međutim, postoje bolesti za koje je genetska predispozicija nastala tijekom procesa evolucije samo kod ljudi. U tim će nam slučajevima komparativna analiza DNK dati vrijedne informacije o genetskoj prirodi takvih bolesti i osjetljivosti ljudi kao vrste na njih.
Analizirajući prikupljene podatke, znanstvenici su napravili svojevrsno računalno preklapanje mape genoma čimpanze na mapu ljudskog genoma, što im je omogućilo identificiranje tri kategorije takozvanih duplikacija DNK - onih koje su prisutne u ljudskom genomu, ali ih nema u genom čimpanze, one koje su prisutne u genomu čimpanze, ali ih nema u ljudskom genomu, te one prisutne u genomima obiju vrsta. Duplikacija DNK je oblik mutacije u kojoj se dio kromosoma udvostručuje. U ovom slučaju uzeti su u obzir segmenti DNA duljine najmanje 20 tisuća parova nukleotida. Ispostavilo se da oko trećine duplikacija DNK pronađenih kod ljudi nema kod čimpanza. Prema Eikleru, ova je brojka iznenadila genetičare jer ukazuje na vrlo visoku učestalost mutacija u kratkom vremenskom razdoblju, prema evolucijskim standardima. U isto vrijeme, analiza duplikacija DNK jedinstvenih za genom čimpanze pokazala je da iako je broj mjesta na kojima se pojavljuju relativno malen, broj kopija dupliciranih segmenata mnogo je veći nego kod ljudi. A u slučajevima kada se udvostručenje DNK događa i kod čimpanza i kod ljudi, kod čimpanza je obično predstavljeno velikim brojem kopija. Konkretno, znanstvenici su otkrili segment koji se pojavljuje 4 puta u ljudskom genomu i 400 puta u genomu čimpanze. Zanimljivo je da se ova regija nalazi u blizini regije koja je kod čimpanza i drugih čovjekolikih majmuna podijeljena na 2 kromosoma, a kod čovjeka je spojena u jedan - kromosom br.2.
No, zapanjujuće razlike između majmuna i ljudi ne objašnjavaju se toliko razlikama u genetskom kodu, koliko različitim aktivnostima gena, naglašava Svante Päbo. Skupina istraživača predvođena njime proučavala je i uspoređivala aktivnost 21 tisuće gena u stanicama srca, jetre, bubrega, testisa i mozga oba primata. Ispostavilo se da ne postoji potpuna podudarnost aktivnosti gena ni u jednom od ovih organa, već su razlike raspoređene krajnje neravnomjerno. Začudo, najmanje razlike znanstvenici su zabilježili u moždanim stanicama - iznosile su svega nekoliko postotaka. A najveće razlike utvrđene su u testisima: ovdje svaki treći gen ima drugačiju aktivnost. No, to je sasvim razumljivo ako se ima u vidu da čimpanze ne formiraju monogamne obitelji, već žive u skupinama, svojevrsnim komunama, koje broje 25-30 jedinki oba spola. Odnosno, "promiskuitet" među čimpanzama mnogo je rašireniji nego među ljudima. Kako bi povećali svoje šanse za razmnožavanje u promiskuitetnim uvjetima, mužjaci čimpanze moraju proizvesti ogromne količine sperme. Nije slučajno da su im testisi deset puta veći od testisa homo sapiensa. Ali ne radi se samo o veličini, naravno, kaže Svante Päbo:
Naši podaci ukazuju na vrlo visoku aktivnost onih gena na Y kromosomu koji su izravno odgovorni za proizvodnju sperme.
A za činjenicu da su ljudi fizički puno slabiji od čimpanza, znanstvenici su pronašli genetsko objašnjenje: kod majmuna mišići rade 5-7 puta učinkovitije jer u svim predstavnicima ljudske rase gen MYH16, koji kodira "miozin" - protein mišićnih vlakana - predstavljen je mutiranom kopijom.
No, ako se koncentriramo na pitanje koja je glavna genetska razlika između čovjeka kao biološke vrste i majmuna i što objašnjava tako uspješno širenje čovjeka tijekom evolucije, onda odgovor, po svemu sudeći, treba tražiti u 6 regija genom identificiran od strane znanstvenika. U ljudskom genomu, ove regije, koje sadrže ukupno nekoliko stotina gena, toliko su stabilne da su praktički identične kod svih ljudi; u genomu čimpanze, naprotiv, često sadrže mutacije. Očigledno, smatraju znanstvenici, ta su područja igrala iznimno važnu ulogu u procesu naše evolucije. Važno je napomenuti da se u jednom od ovih područja nalazi gen FOXP2, jedan od 4 gena odgovorna za razvoj govora. Kao što su eksperimenti pokazali, u laboratorijskim uvjetima majmuni mogu naučiti prilično značajan skup znakova i simbola; čimpanze koje žive u divljini koriste vrlo bogat raspon zvukova za komunikaciju; međutim, fizički nisu u stanju napraviti pokrete usnama i jezikom koji su potrebni za artikulirani govor. Možda je upravo mutacija gena FOXP2 postala jedan od ključnih čimbenika koji su odredili tako različite evolucijske sudbine različitih vrsta primata.
No, ne treba zaboraviti da se čovjek među ostalim životinjskim vrstama nije isticao samo razvijenim govorom. Ali koje su genetske strukture unaprijed odredile uspravno držanje i brzi rast volumena mozga, što je podrazumijevalo sve ostalo, bilo da se radi o stvaranju alata ili korištenju vatre - znanstvenici još ne riskiraju iznijeti hipoteze o tome.
potpuno definiran. Stoga rad na dešifriranju genoma nematoda treba smatrati vrlo uspješnim.
Još veći uspjeh povezan je s dešifriranjem genoma Drosophile, samo u
2 puta manja od ljudske DNK i 20 puta veća od DNK nematode. Unatoč visokom stupnju genetskog poznavanja Drosophile, oko 10% njezinih gena do sada je bilo nepoznato. Ali najparadoksalnije je to što drozofila, koja je mnogo bolje organizirana od nematode, ima manje gena od mikroskopske valjkaste gliste! Sa suvremenog biološkog gledišta to je teško objasniti. Više gena od onih Drosophile prisutno je iu dešifriranom genomu biljke iz obitelji križnica - Arabidopsis, koju genetičari naširoko koriste kao klasični eksperimentalni objekt.
Razvoj genomskih projekata pratio je intenzivan razvoj u mnogim područjima znanosti i tehnologije. Time je bioinformatika dobila snažan poticaj za svoj razvoj. Stvoren je novi matematički aparat za pohranjivanje i obradu ogromnih količina informacija; dizajnirani su sustavi superračunala s neviđenom snagom; Napisane su tisuće programa koji omogućuju, u nekoliko minuta, usporednu analizu različitih blokova informacija, unos novih podataka u računalne baze podataka dnevno,
dobivene u raznim laboratorijima diljem svijeta, te prilagoditi nove informacije onima koje su ranije prikupljene. Istodobno su razvijeni sustavi za učinkovito izdvajanje različitih elemenata genoma i automatsko sekvenciranje, odnosno određivanje nukleotidnih sekvenci DNA. Na temelju toga dizajnirani su snažni roboti koji značajno ubrzavaju sekvenciranje i čine ga jeftinijim.
Razvoj genomike doveo je pak do otkrića ogromnog broja novih činjenica. Značaj mnogih od njih tek treba procijeniti
budućnost. No već sada je očito da će ova otkrića dovesti do preispitivanja mnogih teorijskih stajališta o nastanku i evoluciji različitih oblika života na Zemlji. Oni će pridonijeti boljem razumijevanju molekularnih mehanizama na kojima se temelji funkcioniranje pojedinih stanica i njihovih međudjelovanja; detaljno dekodiranje mnogih još nepoznatih biokemijskih ciklusa;
analiza njihove povezanosti s temeljnim fiziološkim procesima.
Dakle, postoji prijelaz sa strukturne genomike na
funkcionalna, što zauzvrat stvara preduvjete za
istraživanje molekularnih osnova funkcioniranja stanica i organizma u cjelini.
Sada prikupljene informacije bit će predmet unutarnje analize
sljedećih nekoliko desetljeća. Ali svaki sljedeći korak unutra
smjer dešifriranja strukture genoma različitih vrsta, rađa nove tehnologije koje olakšavaju proces dobivanja informacija. Tako,
korištenje podataka o građi i funkciji gena niže organiziranih vrsta živih bića može znatno ubrzati potragu
zamjenjuju prilično radno intenzivne molekularne metode traženja gena.
Najvažnija posljedica dešifriranja strukture genoma određene vrste jest sposobnost identificiranja svih njezinih gena i,
sukladno tome, identifikacija i određivanje molekularne prirode transkribiranih RNA molekula i svih njezinih proteina. Po analogiji s genomom rođeni su koncepti transkriptoma, koji ujedinjuje skup molekula RNA nastalih kao rezultat transkripcije, i iproteoma, koji uključuje mnoge proteine kodirane genima. Tako genomika stvara temelj za intenzivan razvoj novih znanosti – proteomike i transkriptomika. Proteomika je proučavanje strukture i funkcije svakog proteina; analiza proteinskog sastava stanice; određivanje molekularne osnove funkcioniranja pojedine stanice koja je
rezultat koordiniranog rada više stotina proteina, i
proučavanje nastanka fenotipske osobine organizma,
koji proizlaze iz koordiniranog rada milijardi stanica.
Vrlo važni biološki procesi odvijaju se i na razini RNA. Njihova analiza je predmet transkriptomike.
Najveći napori znanstvenika iz mnogih zemalja svijeta koji se bave genomikom bili su usmjereni na rješavanje međunarodnog projekta „Ljudski genom“. Značajan napredak u ovom području vezan je uz realizaciju ideje,
predložio J. S. Venter, pretraži i analiziraj
eksprimirane sekvence DNA, koje se kasnije mogu koristiti kao svojevrsni “tagovi” ili markeri određenih regija genoma. Još jedan samostalan i ništa manje plodan pristup korišten je u radu grupe koju je vodio vlč.
Collins. Temelji se na primarnoj identifikaciji gena za nasljedne ljudske bolesti.
Dekodiranje strukture ljudskog genoma dovelo je do senzacionalnog otkrića. Pokazalo se da ljudski genom sadrži samo 32.000 gena, što je nekoliko puta manje od broja proteina. U isto vrijeme, postoji samo 24 000 gena koji kodiraju proteine; produkti preostalih gena su molekule RNA.
Postotak sličnosti u sekvencama nukleotida DNA između različitih pojedinaca, etničkih skupina i rasa je 99,9%.
Ova sličnost je ono što nas čini ljudima – Homo sapiens! Sva naša varijabilnost na razini nukleotida stane u vrlo skromnu brojku - 0,1%.
Dakle, genetika ne ostavlja mjesta za ideje nacionalne ili rasne superiornosti.
Ali pogledajmo jedni druge – svi smo različiti. Nacionalne, a još više rasne razlike su još uočljivije. Dakle, koji broj mutacija određuje ljudsku varijabilnost, ne u postotku, već u apsolutnom iznosu? Da biste dobili ovu procjenu, morate se sjetiti veličine genoma. Duljina molekule ljudske DNA je
3,2x109 parova baza. 0,1% od toga je 3,2 milijuna nukleotida. No zapamtite da kodirajući dio genoma zauzima manje od 3% ukupne duljine molekule DNA, a mutacije izvan ove regije najčešće nemaju nikakav učinak na fenotipsku varijabilnost. Dakle, da bismo dobili integralnu procjenu broja mutacija koje utječu na fenotip, trebamo uzeti 3% od 3,2 milijuna nukleotida, što će nam dati brojku reda veličine 100 000. To jest, oko 100 tisuća mutacija tvori naš fenotipski varijabilnost. Ako tu brojku usporedimo s ukupnim brojem gena, ispada da u prosjeku postoje 3-4 mutacije po genu.
Koje su to mutacije? Velika većina njih (najmanje 70%)
određuje našu individualnu nepatološku varijabilnost, ono što nas razlikuje, ali nas ne čini lošijima jedne u odnosu na druge. To uključuje karakteristike kao što su boja očiju, kosa, koža, tip tijela, visina, težina,
tip ponašanja koji je također u velikoj mjeri genetski uvjetovan, i još mnogo toga. Oko 5% mutacija povezano je s monogenskim bolestima. Otprilike četvrtina preostalih mutacija pripada klasi funkcionalnih polimorfizama. Oni sudjeluju u formiranju nasljedne predispozicije za raširenu multifaktorijalnu patologiju. Naravno, ove procjene su prilično grube,
ali omogućuju prosuđivanje strukture ljudske nasljedne varijabilnosti.
Poglavlje 1.16. Molekularno genetske osnove evolucije
Revolucija u području molekularne biologije koja se dogodila na prijelazu tisućljeća, a koja je kulminirala dešifriranjem strukture genoma mnogih stotina vrsta mikroorganizama, kao i nekih vrsta protozoa,
kvasci, biljke, životinje i ljudi, preokrenuli su mnoge tradicionalne ideje klasične genetike i približili mogućnost proučavanja molekularnih mehanizama evolucije i specijacije. Rođena je nova znanost - komparativna genomika,
što omogućuje registraciju pojavljivanja u različitim filogenetskim linijama evolucijski značajnih događaja koji se događaju na razini pojedinačnih molekula. Pokazalo se da je u općem slučaju evolucijski napredak povezan ne samo, i ne toliko s povećanjem broja, opsega pa čak i složenosti strukturne organizacije gena, već u mnogo većoj mjeri s promjenom regulacije gena. njihovog rada, koji određuje koordinaciju i tkivnu specifičnost ekspresije desetaka tisuća gena. To je, u konačnici, dovelo do pojave u višim organizmima složenijih, vrlo specifičnih, višenamjenskih kompleksa međusobno povezanih proteina sposobnih za obavljanje fundamentalno novih zadataka.
Razmotrimo prirodu promjena koje se događaju u procesu evolucije na tri informacijske razine: DNA - RNA - protein ili genom - transkriptom - proteom. Općenito, možemo reći da se s povećanjem složenosti organizacije života povećava i veličina genoma. Dakle, veličina DNA prokariota ne prelazi 8x106 bp, postaje dva puta veća kod kvasaca i protozoa, 10-15 puta veća kod insekata, a kod sisavaca porast doseže 3 reda veličine, odnosno tisuću puta (103 ).
Međutim, ova ovisnost nije linearna. Stoga kod sisavaca više ne opažamo značajno povećanje veličine genoma. Osim toga, nije uvijek moguće promatrati odnos između veličine genoma i složenosti organizacije života. Dakle, kod nekih biljaka veličina genoma je red veličine ili čak dva reda veličine veća od one kod ljudi. Podsjetimo, povećanje veličine genoma eukariota u odnosu na prokariote događa se uglavnom zbog pojave nekodirajućih sekvenci, odnosno izbornih elemenata. Već smo rekli da u ljudskom genomu egzoni ne čine više od 1-3%. To znači da broj gena u višim organizmima može biti samo nekoliko puta veći nego u mikroorganizmima.
Sve veća složenost eukariotske organizacije djelomično se objašnjava pojavom dodatnog regulatornog sustava potrebnog za
osiguranje tkivne specifičnosti ekspresije gena. Jedna od posljedica diskontinuirane organizacije gena koja se pojavila kod eukariota bila je raširena pojava alternativnog spajanja i alternativne transkripcije. To je dovelo do pojave novog svojstva u ogromnom broju gena - sposobnosti kodiranja više funkcionalno različitih izoformi proteina. Dakle, ukupna količina proteina
to jest, veličina proteoma; viši mogu imati nekoliko puta veći broj gena.
U prokariota je dopuštena intraspecifična varijabilnost u broju gena, i
slične razlike između različitih sojeva mnogih mikroorganizama, u
uključujući i patogene, može iznositi desetke postotaka. Štoviše, složenost organizacije različitih vrsta mikroorganizama izravno je povezana s brojem i duljinom kodirajućih sekvenci.
Stoga je fenotipska intra- i interspecifična varijabilnost u strogoj vezi s vrlo sličnim veličinama transkriptoma i proteoma. U eukariota je broj gena strogo određena karakteristika vrste, a povećanje evolucijske složenosti temelji se na drugom principu - različitoj višerazinskoj uporabi različitih komponenti ograničenog i prilično stabilnog proteoma.
Sekvenciranje genoma nematoda i Drosophile pokazalo je da su veličine proteoma u ovim vrlo različitim vrstama vrlo slične i samo dvostruko veće od onih kod kvasca i nekih vrsta bakterija. Ovaj obrazac - značajno povećanje složenosti organizacije različitih oblika života uz zadržavanje ili relativno malo povećanje veličine proteoma - karakterističan je za svu kasniju evoluciju do čovjeka. Tako,
Proteomi ljudi i miševa praktički se ne razlikuju jedni od drugih i manje su od 2 puta veći od proteoma mikroskopske nematode valjkaste gliste ili vinske mušice Drosophila. Štoviše, identitet nukleotidnih sekvenci ljudske DNA i
čovjekolikih majmuna iznosi 98,5%, au kodirajućim regijama doseže 99%. Ove se brojke malo razlikuju od vrijednosti od 99,9%,
određivanje intraspecifične sličnosti u sekvencama nukleotida DNA između različitih jedinki, naroda i rasa koje nastanjuju naš planet. Dakle, koje su promjene, koje ne čine više od 1,5% cijelog genoma, ključne za formiranje osobe? Odgovor na ovo pitanje, očito, treba tražiti ne samo na genomskoj i proteomskoj razini.
Doista, uz relativnu stabilnost proteoma, u
U procesu evolucije dolazi do naglog povećanja veličine i složenosti organizacije eukariotskog transkriptoma zbog pojave u genomu ogromnog broja transkribirane i nekodirajuće DNK, kao i značajnog širenja razred gena koji kodiraju RNA. RNA koje ne kodiraju proteine, čiji su glavni izvor introni,
čine veliku većinu transkriptoma viših organizama,
dostižući 97-98% svih transkripcijskih jedinica. Funkcije ovih molekula trenutno se intenzivno analiziraju.
Stoga se ključne evolucijske promjene događaju u pozadini povećanja veličine genoma, prilično stabilnog proteoma i naglog povećanja veličine transkriptoma – sl. 31.
Slika 31. Evolucijske promjene koje se događaju u tri
informacijske razine Pritom je očit prijelaz s jednostavnih oblika života na složenije
korelira s pojavom i širokom distribucijom u genomu dviju temeljnih i donekle međusobno povezanih evolucijskih tekovina: nekodirajuće DNK i ponavljajućih elemenata. Izravna posljedica ovih promjena koje se događaju na genomskoj razini je pojava u procesu evolucije ogromnog broja RNA koje ne kodiraju proteine.
Što je strukturna osnova ovih evolucijskih transformacija?
Svi glavni evolucijski prijelazi: od prokariota do eukariota, od protozoa do metazoa, od prvih životinja do bilateralnih životinja i od primitivnih hordata do kralježnjaka, bili su popraćeni naglim povećanjem složenosti genoma. Očigledno, takvi skokovi u evoluciji rezultat su rijetkih slučajeva uspješnog stapanja cijelih genoma različitih vrsta koje pripadaju sustavnim klasama koje su se znatno udaljile jedna od druge. Tako je simbioza Archaea i Bacteria označila početak prijelaza iz prokariota u eukariote. Očito je da su se kao rezultat endosimbioze pojavili i mitohondriji, kloroplasti i neke druge stanične organele. Temeljno svojstvo viših eukariota, diploidija, nastalo je kao rezultat dobro reguliranog dupliciranja genoma koje se dogodilo prije otprilike 500 milijuna godina.
Umnožavanje genoma unutar vrste događalo se prilično često, i
primjeri za to su brojni slučajevi poliploidije kod biljaka,
gljive pa čak ponekad i kod životinja. Međutim, potencijalni mehanizmi
koji dovode do pojave temeljno novih oblika života u procesu evolucije nisu autopoliploidije, već hibridizacija i horizontalni prijenos ili spajanje genoma. Važno je napomenuti da se najznačajnije evolucijske transformacije, popraćene spajanjem cijelih genoma, događaju u izvanrednim uvjetima, tijekom razdoblja velikih geoloških prijelaza, poput promjena koncentracije kisika u atmosferi, glacijacije Zemlje ili kambrija. Eksplozija.
U relativno mirnim geološkim uvjetima, duplikacije pojedinih gena ili kromosomskih segmenata s njihovom kasnijom divergencijom pokazuju se značajnijima za evoluciju. Usporedba nukleotidnih sekvenci sekvenciranih genoma pokazuje da je učestalost umnožavanja gena prilično visoka i u prosjeku iznosi 0,01 po genu na milijun godina. Velika većina njih se ne manifestira sljedećih nekoliko milijuna godina, i to samo u rijetkim slučajevima
U slučajevima, duplicirani geni mogu steći nove adaptivne funkcije. Međutim, velika klasa "tihih" duplikacija gena služi kao neka vrsta rezervnog fonda za rađanje novih gena i formiranje novih vrsta. Ljudski genom sadrži od 10 do 20 tisuća kopija procesiranih gena koji su nastali retropozicijom mRNA.
Većina njih pripada klasi pseudogena, odnosno ne izražavaju se zbog prisutnosti mutacija ili zbog umetanja u transkripcijski neaktivne regije genoma. Međutim, neki od ovih gena su aktivni, a priroda njihove ekspresije, pa čak i funkcije mogu biti različite,
od onih gena osnivača.
Imaju posebnu ulogu u evoluciji primata i čovjeka. segmentalne duplikacije, koji pripada klasi niske kopije ponavljanja (LCR) i
nastao prije manje od 35 milijuna godina. Ove sekvence su vrlo identični blokovi DNK, koji variraju u veličini od jedne do nekoliko stotina kilobaza. Najčešće su segmentne duplikacije lokalizirane u pericentromernim ili telomernim regijama različitih kromosoma, a ukupno zauzimaju oko 5% ljudskog genoma.
U drugim sekvenciranim genomima nisu pronađene segmentne duplikacije.
Minimalni modul segmentne duplikacije, nazvan duplikon, sadrži fragmente nepovezanih neobrađenih gena, i
to ga razlikuje od drugih poznatih vrsta ponovljenih nizova. Pod određenim uvjetima, duplikoni mogu poslužiti kao izvori za stvaranje novih himernih transkribiranih gena ili obitelji gena iz različitih kombinacija kodirajućih egzona prisutnih u njima. Procjenjuje se da između 150 i 350 gena može razlikovati genome čimpanze i čovjeka.
Ne umanjujući važnost pojave novih i nestanka starih kodnih sekvenci za specijaciju, treba naglasiti stvarnu mogućnost postojanja drugih mehanizama, tj.
igra odlučujuću ulogu u evoluciji eukariota.
Jedan od pokretačkih mehanizama evolucije su pokretni elementi, koji se nalaze u svim vrstama proučavanim u tom smislu.
Genomske promjene koje prate proces specijacije mogu uključivati opsežne reorganizacije kariotipa, lokalne kromosomske preraspodjele, duplikacije obitelji gena, modifikacije pojedinačnih gena,
popraćeno njihovim rođenjem ili gubitkom, kao i razlikama u ekspresiji gena, reguliranim i na razini transkripcije i na razini spajanja ili translacije. Mobilni elementi izravno su povezani sa svim tim procesima.
U nekim slučajevima, prijenosni elementi sami nose sekvence koje kodiraju enzime čija je prisutnost neophodna za transpoziciju DNA ili retropoziciju RNA.
Slične sekvence prisutne su u genomu retrovirusa, LTR-
elemenata i transpozona. Retrotranspozoni također uključuju najbrojniju klasu prijenosnih elemenata - Alu ponavljanja. Po prvi put Alu-
ponavljanja se pojavljuju kod primata prije otprilike 50-60 milijuna godina iz malog gena koji kodira RNA. U procesu daljnje evolucije dolazi do divergencije i snažnog pojačanja ove obitelji. Prijelaz s primata na čovjeka prati eksplozivan porast broja
Alu ponavlja, čiji broj primjeraka, prema nekim procjenama, doseže
1,1 milijun. Alu ponavljanja zauzimaju oko 10% ljudskog genoma, no njihova je distribucija neravnomjerna jer su većinom povezani s genima. Ti su elementi rijetko prisutni u kodirajućim egzonima i prilično se često nalaze u intronima i nekodirajućim regijama mRNA, utječući na stabilnost ovih molekula i/ili učinkovitost translacije. Prisutnost Alu sekvenci u intronskim regijama gena može biti popraćena promjenom u prirodi obrade preRNA, budući da te sekvence sadrže regije homologne donorskim i akceptorskim mjestima spajanja. Kada se Alu elementi umetnu u regulatorne regije gena, transkripcija može biti poremećena, što rezultira
Tim istraživača sa sveučilišta Chicago, Wisconsin i Nebraska-Lincoln opovrgnuo je jednu od klasičnih hipoteza evolucijske prilagodbe na primjeru vinske mušice Drosophila vulgaris (Drosophila). Drosophila melanogaster). U 20. stoljeću te su muhe postale glavni model organizma za razvojnu biologiju i od tada se često koriste u genetskim eksperimentima.
Znanstvenici su sintetizirali drevnu verziju gena jedne vrste i stvorili transgene kukce (to jest, s genima koji se ne mogu steći prirodnim križanjem). Pokusi s njima pokazali su da se prilagodba muha na promjenjive uvjete okoliša tijekom razvoja vrste nije dogodila kako se dosad mislilo. (U nastavku će biti dat konkretan primjer s objašnjenjem.)
"Jedan od glavnih ciljeva moderne evolucijske biologije je utvrditi koji su geni omogućili vrste. Ali to je teško učiniti izravno jer ne možemo testirati kako bismo procijenili utjecaj drevnih gena na životinjsku biologiju", kaže Mo Siddiq sa Sveučilišta u Chicagu. "Shvatili smo da bismo mogli riješiti ovaj problem koristeći dvije nedavno razvijene tehnike - statističku rekonstrukciju drevnih nukleotidnih sekvenci i stvaranje transgenih organizama."
Jednostavno rečeno, biolozi su odlučili najprije "vratiti sat unatrag" kako bi shvatili kakvi su geni vinske mušice bili prije, zatim sintetizirati jedan od njih i umetnuti ga u genom insekta kako bi vidjeli kako će se njegov život promijeniti.
Kada znanstvenici proučavaju prilagodbu na molekularnoj razini, analiza sekvenci nukleotida u DNK pomaže im identificirati takozvane selekcijske potpise, koji ukazuju na brze promjene u genu u prošlosti. No, takvi dokazi zapisani u genomu mogu se smatrati samo posrednim, jer razloga za evoluciju gena može biti puno, a oni nisu nužno povezani s promijenjenim uvjetima okoline na koje se organizam prilagodio. Prema tome, prva metoda koju je gore spomenuo Siddique nije dovoljna.
U novoj studiji znanstvenici su pokušali izravno procijeniti utjecaj evolucije gena na prilagodbu, dodajući drugu metodu prvoj. Voditelj rada, Joe Thornton, u svojim je prethodnim studijama već koristio metodu statističke rekonstrukcije nukleotidnih sekvenci, oslanjajući se na opsežne baze podataka o strukturi genoma modernih organizama, sintetizirao ih i analizirao molekularna svojstva dobivenih gena. u laboratoriju.
Predložio je da bi genetski inženjering i rekonstrukcija drevnih gena zajedno mogli pokazati kako promjene u genomu mogu utjecati na organizam kao cjelinu.
„Ova tehnika stvaranja drevne verzije životinja može se primijeniti na proučavanje različitih aspekata evolucije", napominje Thornton. „Za prvi eksperiment odabrali smo klasičan primjer prilagodbe - vinsku mušicu, koja je tijekom procesa evolucije stekao sposobnost sadržanu u trulom voću.Otkrili smo da je široko rasprostranjena "Hipoteza o mehanizmima evolucije vinskih mušica jednostavno je pogrešna."
Objasnimo što je u divljini D. melanogaster hrani se proizvodima fermentacije voća (često se pojavljuju mali insekti koji se brzo razmnožavaju na mjestima gdje leži pohabano voće). Sposobni su tolerirati veće koncentracije alkohola od svojih najbližih srodnika koji se hrane drugom hranom.
Prije 25 godina, biolozi sa Sveučilišta u Chicagu Martin Kreitman i John McDonald razvili su svoju statističku metodu za identifikaciju selekcijskih potpisa, koja je i danas jedna od najčešće korištenih metoda. Pokazali su njegovu valjanost na primjeru gena alkohol dehidrogenaze (ADH). Ovaj gen kodira enzim koji razgrađuje etanol u stanicama jetre.
Kreitman i McDonald pronašli su tragove selekcije u nukleotidnoj sekvenci ADH, a budući da su znali da tijelo vinske mušice razgrađuje alkohole brže nego njeni srodnici, znanstvenici su sugerirali da je upravo enzim kodiran ADH pomogao mušici da se prilagodi na visoke koncentracije alkohola. Kao rezultat toga, ovaj je rad postao prvi slučaj koji je priznala znanstvena zajednica u kojem je određeni gen utjecao na adaptivnu evoluciju vrste.
Sada su istraživači iz Sjedinjenih Država odlučili testirati hipotezu pomoću drugih tehnologija. Modelirali su varijacije u nukleotidnim sekvencama gena prije i nakon što je vinska mušica stekla toleranciju na etanol (prije oko 2-4 milijuna godina). Znanstvenici su zatim sintetizirali te gene, pokrenuli njihovu ekspresiju i testirali sposobnost dobivenih proteina da razgrade alkohole. Kao rezultat toga, pokazalo se da promjene koje je genom vinske mušice prošao tijekom evolucije nisu imale poseban utjecaj na funkcioniranje enzima.
Zatim su znanstvenici umetnuli "drevni" oblik gena ADH u genom modernih vinskih mušica i uzgojili tisuće modificiranih kukaca kako bi testirali koliko brzo bi razgradili etanol i koliko bi dugo mogli preživjeti ako bi se hranili trulim voćem s visokim udjelom alkohola .
Eksperimenti su pokazali da vinske mušice sa starijim oblikom ADH gena ne obrađuju etanol ništa gore od insekata s "najnovijom verzijom" gena. Štoviše, odrasli su i razmnožavali se na isti način, jedući hranu s visokim udjelom alkohola.
Dakle, nije potvrđena klasična hipoteza, točnije forma D. melanogaster tijekom procesa evolucije doduše prilagodio hrani bogatoj alkoholima, ali promjene u enzimu alkohol dehidrogenaze nisu s time povezane, zaključuju znanstvenici.
Kao što Thornton objašnjava, hipoteza o ADH genu bila je prihvaćena u to vrijeme jer su ekologija, fiziologija i statistički dokazi selekcije upućivali u istom smjeru.
"Ali posredni dokazi ne znače da je pretpostavka nužno točna. Zato smo, sada kada nam je tehnologija dala ovu priliku, htjeli izravno testirati ovu hipotezu", dodaje.
Tim američkih istraživača nada se da će nova tehnika za dobivanje organizama sa starijim verzijama gena postati takozvani zlatni standard u ovom području i da će u budućnosti pomoći da se utvrdi koje su točno genetske promjene utjecale na evolucijske karakteristike organizama.
Rad američkih genetičara detaljnije je opisan u znanstvenom članku objavljenom u časopisu Nature Ecology & Evolution.
Prisjetimo se da smo ranije govorili o tome kako su dubokomorske alge natjerale znanstvenike na evolucijsku prilagodbu. Osim toga, mogućnosti ultrabrze evolucije nedavno je pokazao .
Izdavačka kuća „BINOM. Knowledge Laboratory izdaje knjigu memoara genetičara Craiga Ventera, Life Decipherd. Craig Venter poznat je po svom radu na čitanju i dešifriranju ljudskog genoma. Godine 1992. utemeljio je Institut za istraživanje genoma (TIGR). Venter je 2010. godine stvorio prvi umjetni organizam na svijetu – sintetsku bakteriju Mycoplasma laboratorium. Pozivamo vas da pročitate jedno od poglavlja knjige, u kojem Craig Venter govori o radu od 1999. do 2000. na sekvencioniranju genoma mušice Drosophila.
Naprijed i samo naprijed
Temeljni aspekti nasljeđa pokazali su se, na naše iznenađenje, sasvim jednostavnima, pa je stoga postojala nada da možda priroda i nije toliko nespoznatljiva, a njezina neshvatljivost, koju razni ljudi uvijek iznova proklamiraju, samo je još jedna iluzija, plod našeg neznanja . To nas čini optimistima, jer da je svijet tako složen kao što neki naši prijatelji tvrde, biologija ne bi imala šanse postati egzaktna znanost.
Thomas Hunt Morgan. Fizičke osnove nasljeđa
Mnogi su me ljudi pitali zašto sam od svih živih bića na našem planetu odabrao baš vinsku mušicu; drugi su se pitali zašto nisam odmah prešao na dešifriranje ljudskog genoma. Stvar je u tome da nam je trebala osnova za buduće eksperimente, htjeli smo biti sigurni u ispravnost naše metode prije nego što potrošimo gotovo 100 milijuna dolara na sekvenciranje ljudskog genoma.
Mala vinska mušica odigrala je veliku ulogu u razvoju biologije, posebice genetike. Rod Drosophila uključuje razne muhe - octene, vinske, jabučne, grožđane i voćne - ukupno oko 26 stotina vrsta. Ali izgovorite riječ "drosophila" i svaki će znanstvenik odmah pomisliti na jednu specifičnu vrstu - Drosophilamelanogaster. Budući da se razmnožava brzo i lako, ova sićušna muha služi kao model organizma evolucijskim biolozima. Njime rasvjetljavaju čudo stvaranja – od trenutka oplodnje do nastanka odraslog organizma. Zahvaljujući Drosophili, napravljena su mnoga otkrića, uključujući otkriće gena koji sadrže homeobox koji reguliraju opću strukturu svih živih organizama.
Svaki student genetike upoznat je s eksperimentima na drozofilama koje je izvodio Thomas Hunt Morgan, otac američke genetike. Godine 1910. primijetio je muške mutante s bijelim očima među uobičajenim crvenookim mušicama. Križao je bjelookog mužjaka s crvenookom ženkom i otkrio da su njihovi potomci bili crvenooki: pokazalo se da su bjelooke recesivna osobina, a sada znamo da su vam potrebne dvije kopije da bi muhe imale bijele oči. gena bjelookih, po jedan od svakog roditelja. Nastavljajući križati mutante, Morgan je otkrio da samo mužjaci pokazuju svojstvo bijelih očiju i zaključio da je to svojstvo povezano sa spolnim kromosomom (Y kromosom). Morgan i njegovi studenti proučavali su nasljedne osobine u tisućama vinskih mušica. Danas se eksperimenti s Drosophilom provode u laboratorijima molekularne biologije diljem svijeta, gdje više od pet tisuća ljudi proučava ovaj mali kukac.
Naučio sam iz prve ruke važnost Drosophile kada sam koristio biblioteke njenih cDNA gena za proučavanje adrenalinskih receptora i otkrio njihov ekvivalent u mušici - oktopaminske receptore. Ovo otkriće ukazalo je na zajedništvo evolucijske nasljednosti živčanog sustava muhe i čovjeka. Pokušavajući shvatiti cDNA biblioteke ljudskog mozga, pronašao sam gene sa sličnim funkcijama kompjutorskom usporedbom ljudskih gena s genima Drosophile.
Projekt sekvenciranja gena Drosophile započeo je 1991. godine, kada su Jerry Rubin s kalifornijskog sveučilišta Berkeley i Allen Spradling s Carnegie instituta odlučili da je vrijeme da preuzmu taj zadatak. Do svibnja 1998. već je bilo dovršeno 25% sekvenciranja, a ja sam dao prijedlog za koji je Rubin rekao da je "predobar da bi se propustio". Moja ideja bila je prilično riskantna: tisuće istraživača vinske mušice iz različitih zemalja moralo bi pomno ispitati svako slovo šifre koju smo dobili, uspoređujući ga s Jerryjevim visokokvalitetnim, referentnim podacima, a zatim donijeti zaključak o prikladnosti moje metode .
Izvorni plan bio je dovršiti sekvenciranje genoma muhe u roku od šest mjeseci do travnja 1999., a zatim započeti napad na ljudski genom. Činilo mi se da je to najučinkovitiji i najjasniji način da pokažemo da naša nova metoda djeluje. A ako ne uspijemo, pomislio sam, onda bi bilo bolje to brzo provjeriti na primjeru Drosophile nego radom na ljudskom genomu. Ali zapravo bi potpuni neuspjeh bio najspektakularniji neuspjeh u povijesti biologije. Jerry je također stavljao svoj ugled na kocku, pa su svi u Celeri bili odlučni podržati ga. Zamolio sam Marka Adamsa da vodi naš dio projekta, a budući da je i Jerry imao vrhunski tim na Berkeleyu, naša je suradnja tekla bez problema.
Prije svega, postavilo se pitanje čistoće DNK koju smo morali sekvencirati. Kao i ljudi, muhe se razlikuju na genetskoj razini. Ako postoji više od 2% genetske varijacije u populaciji, a imamo 50 različitih jedinki u odabranoj skupini, tada se dekodiranje pokazuje vrlo teškim. Jerryjev prvi korak bio je ukrštanje muha što je više moguće kako bismo dobili jedinstvenu varijantu DNK. Ali parenje u srodstvu nije bilo dovoljno da se osigura genetska čistoća: prilikom izdvajanja DNK muhe, postojao je rizik od kontaminacije genetskim materijalom iz bakterijskih stanica u hrani muhe ili u njezinim crijevima. Kako bi izbjegao te probleme, Jerry je radije izdvojio DNK iz embrija muhe. Ali čak i iz embrionalnih stanica, prvo smo morali izolirati jezgre s potrebnom DNK, kako je ne bi kontaminirali ekstranuklearnom DNK mitohondrija - "elektrane" stanice. Kao rezultat, dobili smo epruvetu s mutnom otopinom čiste DNA Drosophile.
U ljeto 1998., Hamov tim, koji je imao tako čist DNK muhe, počeo je stvarati biblioteke njegovih fragmenata. Sam Ham najviše je volio rezati DNK i preklapati dobivene fragmente, smanjujući osjetljivost svog slušnog aparata kako ga nikakvi strani zvukovi ne bi ometali u poslu. Stvaranje knjižnica trebalo je biti početak velikog sekvenciranja, ali do sada su se posvuda čuli samo zvukovi bušilica, čekića i pila. Cijela armija graditelja stalno je bila trn u oku u blizini, a mi smo nastavili rješavati najvažnije probleme - rješavanje problema u radu sekvencera, robota i druge opreme, pokušavajući ne u godinama, već u nekoliko mjeseci stvoriti pravu tvornicu sekvenciranja " od nule.
Prvi model 3700 DNK sekvencer isporučen je Celeri 8. prosinca 1998. uz veliko uzbuđenje i zajednički uzdah olakšanja. Uređaj je izvađen iz drvene kutije, smješten u prostoriju bez prozora u podrumu - svoj privremeni dom, i odmah započelo testiranje. Nakon što je počelo djelovati, dobili smo vrlo kvalitetne rezultate. Ali ti rani sekvenceri bili su prilično nestabilni, a neki su bili neispravni od samog početka. Bilo je i stalnih problema s radnicima, ponekad gotovo svakodnevnih. Na primjer, pojavila se ozbiljna pogreška u programu za upravljanje robotskim manipulatorom - ponekad se mehanička ruka robota ispružila preko uređaja velikom brzinom i zabila se u zid. Kao rezultat toga, sekvencer je stao i morao je biti pozvan tim za popravak da ga popravi. Neki sekvenceri nisu uspjeli zbog zalutale laserske zrake. Za zaštitu od pregrijavanja korištene su folija i traka jer su pri visokim temperaturama žuto obojeni Gs fragmenti isparili iz sekvenci.
Iako su se uređaji sada isporučivali redovito, oko 90% njih bilo je neispravno od samog početka. Nekih dana sekvenceri uopće nisu radili. Čvrsto sam vjerovao u Mikea Hunkapillera, ali moja je vjera bila jako poljuljana kada je za naše neuspjehe počeo kriviti naše zaposlenike, građevinsku prašinu, najmanje kolebanje temperature, mjesečeve mijene i tako dalje. Neki od nas su i posijedjeli od stresa.
Mrtvi 3700-i sjedili su u kantini čekajući da ih pošalju natrag u ABI, i na kraju je došlo do točke da smo morali ručati praktički u mrtvačnici sekvencera. Bio sam u očaju - nakon svega, trebao mi je određeni broj radnih uređaja svaki dan, točnije 230! Za oko 70 milijuna dolara ABI nam je obećao osigurati ili 230 savršeno funkcionalnih uređaja koji bi radili cijeli dan bez prekida ili 460 koji bi radili barem pola dana. Osim toga, Mike je trebao udvostručiti broj kvalificiranog tehničkog osoblja da odmah popravi sekvencer nakon kvara.
No, kakav je interes sve to raditi za isti novac! Osim toga, Mike sada ima još jednog klijenta - vladin genomski projekt, čiji su čelnici već počeli kupovati stotine uređaja bez ikakvog testiranja. Budućnost Celere ovisila je o tim sekvencerima, ali Mike očito nije shvaćao da budućnost ABI-ja također ovisi o njima. Sukob je bio neizbježan, kao što je bilo očito na važnom sastanku između ABI inženjera i mog tima u Celeri.
Nakon što smo izvijestili o ogromnom broju neispravnih instrumenata i koliko je dugo trebalo da se poprave kvarovi na sekvenceru, Mike je opet pokušao svu krivnju svaliti na moje zaposlenike, ali čak se ni njegovi inženjeri nisu složili s njim. Tony White je na kraju intervenirao. "Nije me briga koliko to košta ili tko treba biti ubijen zbog toga", rekao je. Tada je prvi i posljednji put stvarno stao na moju stranu. Naredio je Mikeu da osigura isporuku novih sekvencera što je brže moguće, čak i na račun drugih kupaca i čak iako se još nije znalo koliko će koštati.
Tony je također naredio Mikeu da angažira još dvadeset tehničara koji će brzo popraviti i utvrditi uzrok svih problema. U stvarnosti je to bilo lakše reći nego učiniti jer je nedostajalo iskusnih radnika. Za početak, Eric Lander je pokrao dva najkvalificiranija inženjera, a po Mikeovom mišljenju, i mi smo bili krivi. Okrećući se Marku Adamsu, Mike je rekao: "Trebali ste ih unajmiti prije nego što je to učinio netko drugi." Nakon takve izjave potpuno sam izgubio svaki respekt prema njemu. Uostalom, prema našem dogovoru, ja nisam mogao zaposliti zaposlenike ABI-ja, dok su Lander i drugi voditelji vladinog projekta genoma imali pravo na to, tako da su vrlo brzo najbolji inženjeri ABI-ja počeli raditi za našu konkurenciju. Do kraja sastanka shvatio sam da problemi ostaju, ali je sinuo tračak nade za poboljšanje.
Tako se i dogodilo, iako ne odmah. Naš arsenal sekvencera povećao se s 230 na 300 uređaja, a ako je 20-25% njih otkazalo, i dalje smo imali oko 200 radnih sekvencera i nekako smo se nosili sa zadacima. Tehničko osoblje radilo je herojski i stalno povećavalo tempo popravaka, smanjujući zastoje. Cijelo to vrijeme mislio sam na jedno: ovo što radimo je izvedivo. Neuspjesi su se događali iz tisuću razloga, ali neuspjeh nije bio dio mojih planova.
Ozbiljno smo počeli sekvencirati genom Drosophile 8. travnja, otprilike u vrijeme kad smo trebali završiti ovaj posao. Ja sam, naravno, razumio da me se White želi riješiti, ali sam učinio sve što je bilo u mojoj moći da izvršim glavni zadatak. Napetost i tjeskoba su me proganjali kod kuće, ali o tim problemima nisam mogao razgovarati sa svojom “osobom od povjerenja”. Claire je pokazala svoj prezir kad je vidjela koliko sam zaokupljen Celerinim poslovima. Osjećala se kao da ponavljam iste greške koje sam radio radeći u TIGR-u/HGS-u. Do 1. srpnja osjećao sam se duboko depresivno, baš kao u Vijetnamu.
Budući da nam pokretna metoda još nije uspjela, morali smo obaviti naporan, iscrpljujući posao - ponovno "zalijepiti" fragmente genoma. Kako bi otkrio podudarnosti bez da ga ometaju ponavljanja, Gene Myers predložio je algoritam temeljen na ključnom principu moje verzije metode sačmarice: sekvenciranje oba kraja svih rezultirajućih klonova. Budući da je Ham dobivao klonove tri točno poznate veličine, znali smo da su dvije terminalne sekvence bile na strogo određenoj udaljenosti jedna od druge. Kao i prije, ova metoda "usparivanja" pružit će nam izvrsnu priliku za ponovno sastavljanje genoma.
Ali budući da je svaki kraj sekvence zasebno sekvenciran, da bi ova metoda sastavljanja točno funkcionirala, bilo je potrebno pažljivo voditi evidenciju - kako bismo bili apsolutno sigurni da smo uspjeli ispravno povezati sve parove krajnjih sekvenci: na kraju krajeva, ako barem jedan od stotinu pokušaja dovodi do pogreške i nije pronađen nijedan odgovarajući par za dosljednost, sve će otići u vodu i metoda neće raditi. Jedan od načina da se to izbjegne je korištenje crtičnih kodova i senzora za praćenje svakog koraka procesa. No, na početku rada laboratorijski tehničari nisu imali potreban softver i opremu za sekvenciranje, pa su sve morali raditi ručno. U Celeri je mali tim od manje od dvadeset ljudi svaki dan obrađivao rekordnih 200.000 klonova. Mogli bismo predvidjeti neke pogreške, kao što je pogrešno očitavanje podataka iz 384 bušotine, a zatim upotrijebiti računalo da pronađemo očigledno pogrešnu operaciju i ispravimo situaciju. Naravno, bilo je još nekih nedostataka, ali to je samo potvrdilo umijeće i uvjerenje ekipe da možemo otkloniti pogreške.
Unatoč svim poteškoćama, uspjeli smo u četiri mjeseca pročitati 3156 milijuna sekvenci, ukupno oko 1,76 milijardi parova nukleotida sadržanih između krajeva 1,51 milijuna DNK klonova. Sada je došao red na Genea Myersa, njegov tim i naše računalo - bilo je potrebno spojiti sve dijelove u kromosome Drosophile. Što su dijelovi postajali dulji, to je slijed postajao manje točan. U slučaju Drosophile, nizovi su u prosjeku imali 551 par baza, a prosječna točnost bila je 99,5%. S obzirom na nizove od 500 slova, gotovo svatko može locirati podudaranja pomicanjem jednog niza duž drugog dok se ne pronađe podudaranje.
Da bismo sekvencirali Haemophilus influenzae, imali smo 26 tisuća sekvenci. Za usporedbu svake od njih sa svim ostalima bilo bi potrebno 26 tisuća usporedbi na kvadrat, odnosno 676 milijuna. Genom Drosophile, sa svojih 3,156 milijuna očitavanja, zahtijevao bi oko 9,9 trilijuna usporedbi. U slučaju čovjeka i miša, gdje smo proizveli 26 milijuna čitanja sekvenci, bilo je potrebno oko 680 trilijuna usporedbi. Stoga ne čudi da je većina znanstvenika bila vrlo skeptična u pogledu mogućeg uspjeha ove metode.
Iako je Myers obećao da će sve popraviti, stalno je imao sumnje. Sada je radio dane i noći, izgledao iscrpljeno i nekako sivo. Osim toga, imao je problema u obitelji, a slobodno vrijeme počeo je provoditi najviše s novinarom Jamesom Shreveom koji je pisao o našem projektu i poput sjene pratio tijek istraživanja. Pokušavajući nekako odvratiti Genea, poveo sam ga sa sobom na Karibe da se opusti i plovi svojom jahtom. Ali čak je i ondje sjedio satima, pogrbljen nad svojim prijenosnim računalom, namrštivši crne obrve i škiljeći svoje crne oči od jarkog sunca. I, unatoč nevjerojatnim poteškoćama, Gene i njegov tim uspjeli su generirati više od pola milijuna linija računalnog koda za novi asembler u šest mjeseci.
Kad bi rezultati sekvenciranja bili 100% točni, bez duplikata DNK, sastavljanje genoma bilo bi relativno jednostavan zadatak. Ali u stvarnosti, genomi sadrže veliki broj ponovljenih DNK različitih tipova, duljina i učestalosti. S kratkim ponavljanjima od manje od pet stotina parova baza relativno je lako izaći na kraj; duža ponavljanja su teža. Kako bismo riješili ovaj problem, upotrijebili smo metodu “pronalaženja para”, odnosno sekvencirali smo oba kraja svakog klona i dobili klonove različitih duljina kako bismo osigurali maksimalan broj podudarnosti.
Algoritmi, kodirani u pola milijuna redaka računalnog koda Jinovog tima, predložili su scenarij korak po korak - od "najbezazlenijih" radnji, poput jednostavnog preklapanja dviju sekvenci, do složenijih, poput korištenja otkrivenih parova za spajanje otoka sekvenci koje se preklapaju. Bilo je to poput slaganja slagalice, gdje se mali otoci sklopljenih dijelova sastavljaju u veće otoke, a zatim se cijeli proces ponovno ponavlja. Samo je naša slagalica imala 27 milijuna dijelova. I bilo je vrlo važno da su dijelovi uzeti iz niza visokokvalitetnog sastavljanja: zamislite što bi se dogodilo ako sastavite slagalicu, a boje ili slike njezinih elemenata budu nejasne i mutne. Za dalekosežni poredak sekvence genoma, značajan udio očitanja mora biti u obliku podudarnih parova. S obzirom na to da su se rezultati još uvijek pratili ručno, s olakšanjem smo otkrili da je 70% sekvenci koje smo imali bilo upravo ovakvo. Računalni modelari objasnili su da bi s nižim postotkom bilo nemoguće sastaviti našeg "Humpty Dumptyja".
A sada smo mogli upotrijebiti Celera asembler za sekvenciranje niza: u prvoj fazi, rezultati su prilagođeni kako bi se postigla najveća točnost; u drugom koraku, program Screener uklonio je kontaminirajuće sekvence iz plazmida ili E. coli DNA. Proces sastavljanja može biti poremećen sa samo 10 parova baza "stranog" niza. U trećem koraku, program Screener provjerio je usklađenost svakog fragmenta s poznatim sekvencama koje se ponavljaju u genomu vinske mušice - podatke je dao Jerry Rubin, koji nam ih je "ljubazno" dostavio. Snimljene su lokacije ponavljanja s djelomično preklapajućim regijama. U četvrtom koraku, drugi program (Overlapper) otkrio je preklapajuća područja uspoređujući svaki fragment sa svim ostalima - kolosalan eksperiment u obradi ogromne količine numeričkih podataka. Uspoređivali smo 32 milijuna fragmenata svake sekunde, s ciljem da pronađemo najmanje 40 parova baza koji se preklapaju s manje od 6% razlika. Kad smo otkrili dvije preklapajuće regije, spojili smo ih u veći fragment, takozvani “contig” - skup preklapajućih fragmenata.
U idealnom slučaju, to bi bilo dovoljno za sastavljanje genoma. Ali morali smo se boriti s zastajkivanjima i ponavljanjima u DNK kodu, što je značilo da se jedan dio DNK može preklapati s nekoliko različitih regija, stvarajući lažne veze. Kako bismo pojednostavili zadatak, ostavili smo samo jedinstveno povezane fragmente, takozvane “unitigove”. Program koji smo koristili za izvođenje ove operacije (Unitigger) u biti je uklonio sve sekvence DNK koje nismo mogli sa sigurnošću identificirati, ostavljajući samo ove jedinice. Ovaj korak ne samo da nam je dao priliku da razmotrimo druge mogućnosti sastavljanja fragmenata, već je i značajno pojednostavio zadatak. Nakon redukcije, broj fragmenata koji se preklapaju smanjen je s 212 milijuna na 3,1 milijun, a problem je pojednostavljen 68 puta. Dijelovi slagalice postupno su ali postojano sjedali na svoje mjesto.
A onda bismo mogli koristiti informacije o načinu na koji su sekvence istog klona uparene pomoću algoritma "kostura". Sve moguće jedinice s međusobno preklapajućim parovima baza kombinirane su u posebne okvire. Kako bih opisao ovu fazu u svojim predavanjima, povlačim analogiju s dječjim setom igračaka za konstrukciju Tinkertoys. Sastoji se od štapića različitih duljina, koji se mogu umetnuti u rupe koje se nalaze na drvenim ključnim dijelovima (kuglama i diskovima) i tako stvoriti trodimenzionalnu strukturu. U našem slučaju, ključni dijelovi su jedinice. Znajući da se uparene sekvence nalaze na krajevima klonova dugih 2 tisuće, 10 tisuća ili 50 tisuća parova baza - to jest, čini se da su na udaljenosti određenog broja rupa jedna od druge - mogu se poredati.
Testiranje ove tehnike na sekvenci Jerryja Rubina, koja je bila otprilike jedna petina genoma vinske mušice, rezultiralo je sa samo 500 praznina. Testirajući na vlastitim podacima u kolovozu, dobili smo više od 800.000 malih fragmenata. Značajno veća količina podataka za obradu pokazala je da je tehnika loše funkcionirala – rezultat je bio suprotan od očekivanog. Sljedećih nekoliko dana panika je rasla, a popis mogućih pogrešaka se produžio. S najvišeg kata zgrade br. 2, navala adrenalina prodrla je u prostoriju koja se u šali naziva "Tranquil Chambers". No, tu nije bilo mira ni spokoja, pogotovo barem nekoliko tjedana, kada su zaposlenici doslovno lutali u krug tražeći izlaz iz situacije.
Problem je na kraju riješio Arthur Delcher, koji je radio s programom Overlapper. Primijetio je nešto čudno na retku 678 od 150.000 redaka koda, gdje je manja razlika značila da važan dio utakmice nije snimljen. Pogreška je ispravljena i 7. rujna imali smo 134 stanične skele koje pokrivaju stvarni (eukromatski) genom vinske mušice. Bili smo oduševljeni i odahnuli. Došao je trenutak da svoj uspjeh objavimo cijelom svijetu.
Konferencija o sekvenciranju genoma, čiji sam domaćin prije nekoliko godina, pružila je izvrsnu priliku za to. Bio sam siguran da će biti veliki broj ljudi željnih da se uvjere jesmo li održali obećanje. Odlučio sam da Mark Adams, Gene Myers i Jerry Rubin govore o našim postignućima, a prije svega o procesu sekvenciranja, sastavljanju genoma i značaju toga za znanost. Zbog navale ljudi koji su htjeli doći na konferenciju, morao sam je preseliti iz Hilton Heada u veći hotel Fontainebleau u Miamiju. Na konferenciji su sudjelovali predstavnici velikih farmaceutskih i biotehnoloških kompanija, stručnjaci za genomska istraživanja iz cijelog svijeta, dosta kolumnista, novinara i predstavnika investicijskih kompanija - svi su bili prisutni. Naši konkurenti iz Incytea potrošili su mnogo novca na organizaciju domjenaka nakon konferencije, korporativno video snimanje itd. - učinili su sve kako bi uvjerili javnost da nude “najdetaljnije informacije o ljudskom genomu”.
Okupili smo se u velikoj konferencijskoj dvorani. Uređena u neutralnim bojama, ukrašena zidnim lampama, bila je predviđena za dvije tisuće ljudi, no ljudi su dolazili i ubrzo je dvorana bila puna do posljednjeg mjesta. Konferencija je otvorena 17. rujna 1999. prezentacijama Jerryja, Marka i Genea na prvoj sjednici. Nakon kratkog uvoda, Jerry Rubin najavio je da će publika čuti o najboljem zajedničkom projektu poznatih tvrtki u kojem je ikad sudjelovao. Atmosfera se zahuktavala. Publika je shvatila da ne bi tako pompozno govorio da nismo pripremili nešto uistinu senzacionalno.
U tišini koja je uslijedila, Mark Adams je počeo detaljno opisivati rad naše "tvorničke radnje" u Celeri i naše nove metode sekvenciranja genoma. No, o okupljenom genomu nije rekao ni riječi, kao da zafrkava publiku. Zatim je Gene izašao i govorio o principima metode sačmarice, o sekvenciranju Haemophilusa i o glavnim fazama asemblera. Pomoću računalne animacije demonstrirao je cijeli proces obrnutog sklapanja genoma. Vrijeme predviđeno za prezentacije je istjecalo, a mnogi su već odlučili da će se sve ograničiti na elementarnu prezentaciju u PowerPointu, bez prezentiranja konkretnih rezultata. Ali onda je Gene sa zlobnim osmijehom primijetio da bi publika vjerojatno ipak željela vidjeti prave rezultate i da se ne bi zadovoljila imitacijom.
Bilo je nemoguće predstaviti naše rezultate jasnije i izražajnije nego što je to učinio Gene Myers. Shvatio je da sami rezultati sekvenciranja neće ostaviti pravi dojam, pa ih je, da bi bio uvjerljiviji, usporedio s rezultatima Jerryjeva mukotrpnog istraživanja tradicionalnom metodom. Ispostavilo se da su identične! Stoga je Jin usporedio rezultate našeg sklopa genoma sa svim poznatim markerima preslikanim na genom vinske mušice prije nekoliko desetljeća. Od tisuće markera samo šest nije odgovaralo rezultatima naše skupštine. Pažljivim ispitivanjem svih šest, uvjerili smo se da je Celerino sekvenciranje ispravno i da su pogreške sadržane u radu obavljenom u drugim laboratorijima korištenjem starih metoda. Konačno, Gene je rekao da smo tek počeli sekvencirati ljudsku DNK i da bi ponavljanja vjerojatno bila manji problem nego kod Drosophile.
Uslijedio je glasan i dugotrajan pljesak. Huk koji nije prestajao u pauzi značio je da smo ostvarili cilj. Jedan od novinara primijetio je kako sudionik vladinog projekta genoma tužno odmahuje glavom: “Čini se da će ovi nitkovi stvarno sve učiniti.” 1 Konferenciju smo napustili s novim nabojem energije.
Ostala su dva važna problema za riješiti, a oba su nam bila poznata. Prvo je kako objaviti rezultate. Unatoč memorandumu o razumijevanju koji smo potpisali s Jerryjem Rubinom, našem poslovnom timu nije odgovarala ideja o prijenosu vrijednih rezultata sekvenciranja Drosophile u GenBank. Predložili su stavljanje rezultata sekvenciranja vinske mušice u zasebnu bazu podataka u Nacionalnom centru za biotehnološke informacije, gdje bi ih svatko mogao koristiti pod jednim uvjetom - ne u komercijalne svrhe. Nabrijani, pušač Michael Ashburner s Europskog instituta za bioinformatiku bio je krajnje nesretan zbog toga. Vjerovao je da je Celera "prevarila sve" 2. (Pisao je Rubinu: "Što se dovraga događa u Celeri?" 3) Collins je također bio nesretan, ali što je još važnije, bio je i Jerry Rubin. Na kraju sam ipak poslao naše rezultate GenBank-u.
Drugi problem ticao se Drosophile - imali smo rezultate sekvenciranja njezinog genoma, ali nismo uopće razumjeli što oni znače. Morali smo ih analizirati ako smo htjeli napisati rad, baš kao što smo prije četiri godine učinili s Haemophilusom. Analiza i karakterizacija genoma muhe mogla bi potrajati više od godinu dana - a ja nisam imao to vrijeme jer sam se sada morao usredotočiti na ljudski genom. Nakon rasprave o tome s Jerryjem i Markom, odlučili smo uključiti znanstvenu zajednicu u rad na Drosophili, pretvarajući ga u uzbudljiv znanstveni problem, i tako brzo pokrenuti stvar naprijed, napravivši zabavan odmor od dosadnog procesa opisa genoma - poput međunarodnog skautskog jamboreea. Nazvali smo to Genomski Jamboree i pozvali vodeće znanstvenike iz cijelog svijeta da dođu u Rockville na otprilike tjedan ili deset dana kako bi analizirali genom muhe. Na temelju dobivenih rezultata planirali smo napisati seriju članaka.
Svima se svidjela ideja. Jerry je počeo slati pozivnice za naš događaj skupinama vodećih istraživača, a Celera stručnjaci za bioinformatiku odlučili su koja će računala i programi biti potrebni kako bi rad znanstvenika bio što učinkovitiji. Dogovorili smo se da im Celera plati troškove puta i smještaja. Među pozvanima su bili i moji najoštriji kritičari, ali smo se nadali da njihove političke ambicije neće utjecati na uspjeh našeg pothvata.
U studenom nam je stiglo oko 40 stručnjaka za drozofile, a čak je i za naše neprijatelje ponuda bila previše primamljiva da bi je odbili. U početku, kada su sudionici shvatili da moraju analizirati više od sto milijuna parova baza genetskog koda u nekoliko dana, situacija je bila prilično napeta. Dok su novopridošli znanstvenici spavali, moje je osoblje radilo danonoćno, razvijajući programe za rješavanje nepredviđenih problema. Do kraja trećeg dana, kada se pokazalo da novi softverski alati omogućuju znanstvenicima, kako je rekao jedan naš gost, “da u nekoliko sati dođu do nevjerojatnih otkrića za koja je prije bio potreban gotovo cijeli život”, situacija se smirila. Svaki dan usred dana, na znak kineskog gonga, svi su se okupljali kako bi razgovarali o najnovijim rezultatima, rješavali tekuće probleme i sastavljali plan rada za sljedeći krug.
Svakim danom rasprave su postajale sve zanimljivije. Zahvaljujući Celeri naši su gosti imali priliku prvi zaviriti u novi svijet, a ono što je otkriveno nadmašilo je očekivanja. Ubrzo se pokazalo da nemamo dovoljno vremena razgovarati o svemu što želimo i shvatiti što to sve znači. Mark je priredio svečanu večeru, koja nije dugo trajala jer su svi brzo požurili natrag u laboratorije. Ubrzo su se ručkovi i večere konzumirali ispred računalnih ekrana na kojima su bili prikazani podaci o genomu Drosophile. Po prvi put otkrivene su dugo očekivane obitelji receptorskih gena, zajedno s iznenađujućim brojem gena vinskih mušica sličnih genima ljudskih bolesti. Svako otkriće popraćeno je radosnim urlicima, zvižducima i prijateljskim tapšanjem po ramenu. Začudo, usred naše znanstvene gozbe jedan je par našao vremena za zaruke.
Bilo je, međutim, zabrinutosti: tijekom rada znanstvenici su otkrili samo oko 13 tisuća gena umjesto očekivanih 20 tisuća. Budući da “niski” crv C. elegans ima oko 20 tisuća gena, mnogi su smatrali da ih vinska mušica mora imati više, budući da ima 10 puta više stanica, pa čak i živčani sustav. Postojao je jedan jednostavan način da se uvjerimo da nema pogreške u izračunima: uzmite 2500 poznatih gena muhe i pogledajte koliko njih možemo pronaći u našem nizu. Nakon pomne analize, Michael Cherry sa Sveučilišta Stanford izvijestio je da je pronašao sve osim šest gena. Nakon rasprave, ovih je šest gena klasificirano kao artefakti. Činjenica da su geni identificirani bez grešaka inspirirala nas je i ulila nam povjerenje. Zajednica tisuća znanstvenika posvećenih istraživanju Drosophile provela je desetljeća prateći tih 2500 gena, a sada ih je čak 13600 bilo pred njima na ekranu računala.
Tijekom neizbježnog fotografiranja na kraju posla, dogodio se nezaboravan trenutak: nakon tradicionalnog tapšanja po ramenu i prijateljskog rukovanja, Mike Ashburner se spustio na sve četiri kako bih se ja ovjekovječila na fotografiji s nogom na njegovim leđima . Stoga je želio – unatoč svim svojim sumnjama i skepticizmu – odati priznanje našim postignućima. Poznati genetičar i istraživač drozofila, čak je osmislio i odgovarajući naslov za fotografiju: "Stoji na ramenima diva." (Imao je prilično krhku figuru.) “Odajmo priznanje onima koji ga zaslužuju”, kasnije je napisao 4 . Naši protivnici pokušali su prikazati kašnjenja u prijenosu rezultata sekvenciranja u javnu bazu podataka kao odstupanje od naših obećanja, ali su i oni bili prisiljeni priznati da je sastanak dao “izuzetno vrijedan doprinos globalnom istraživanju vinske mušice” 5 . Nakon što su iskusili što je prava “znanstvena nirvana”, svi su se rastali kao prijatelji.
Odlučili smo objaviti tri velika rada: jedan o sekvenciranju cijelog genoma s Mikeom kao prvim autorom, jedan o sastavljanju genoma s Geneom kao prvim autorom i treći o komparativnoj genomici crva, kvasca i ljudskog genoma s Jerryjem kao prvim autorom. Autor. Radovi su predani Scienceu u veljači 2000. i objavljeni u posebnom broju 24. ožujka 2000., manje od godinu dana nakon mog razgovora s Jerryjem Rubinom u Cold Spring Harboru. 6 Prije objavljivanja, Jerry je organizirao da govorim na godišnjoj konferenciji o istraživanju Drosophile u Pittsburghu, na kojoj su sudjelovale stotine najeminentnijih ljudi na tom području. Na svaku stolicu u prostoriji moje je osoblje stavilo CD s cijelim genomom Drosophile, kao i reprinte naših radova objavljenih u Scienceu. Jerry me vrlo srdačno predstavio, uvjeravajući okupljene da sam ispunio sve svoje obveze i da smo dobro surađivali. Moje predavanje završilo je izvješćem o nekim istraživanjima provedenim tijekom sastanka i kratkim komentarom podataka na CD-u. Pljesak nakon mog govora bio je jednako iznenađujući i ugodan kao i prije pet godina kada smo Ham i ja prvi put predstavili genom Haemophilusa na mikrobiološkoj konvenciji. Nakon toga, radovi o genomu Drosophile postali su najčešće citirani radovi u povijesti znanosti.
Iako su tisuće istraživača vinskih mušica diljem svijeta bile oduševljene rezultatima, moji su kritičari brzo krenuli u ofenzivu. John Sulston nazvao je pokušaj sekvenciranja genoma muhe neuspješnim, iako je sekvenca koju smo dobili bila potpunija i točnija od rezultata njegovog mukotrpnog desetogodišnjeg napora da sekvencira genom crva, za koji je trebalo još četiri godine nakon objavljivanja nacrta u časopisu Science. Sulstonov kolega Maynard Olson nazvao je sekvencu genoma Drosophile "sramotom" koju će vladin Projekt ljudskog genoma morati riješiti, "milošću" Celere. Zapravo, tim Jerryja Rubina uspio je brzo zatvoriti preostale praznine u sekvenci objavljivanjem i komparativnom analizom već sekvenciranog genoma u manje od dvije godine. Ovi podaci potvrđuju da smo imali 1-2 pogreške na 10 kb u cijelom genomu i manje od 1 pogreške na 50 kb u radnom (eukromatskom) genomu.
Međutim, unatoč općem hvaljenju projekta Drosophila, napetosti u mom odnosu s Tonyjem Whiteom dosegle su vrhunac u ljeto 1999. godine. White se nije mogao pomiriti s pažnjom koju je tisak posvetio mojoj osobi. Svaki put kad je dolazio u Celeru, prolazio je pokraj primjeraka članaka o našim uspjesima koji su visjeli na zidovima u hodniku pokraj mog ureda. A ovdje smo uvećali jednu od njih - naslovnicu nedjeljnog priloga lista USA Today. Na njemu pod naslovom “Hoće li ovaj AVANTURIST doći do najvećeg znanstvenog otkrića našeg vremena?” 7 pokazao me, u plavoj kariranoj košulji, kako prekrižim noge, a oko mene Kopernik, Galileo, Newton i Einstein lebde u zraku - a od Whitea ni traga.
Svakoga dana njegova je tajnica za tisak zvala da vidi može li Tony sudjelovati u naizgled beskrajnom nizu intervjua koji su se odvijali u Celeri. Malo se smirio – i to samo nakratko, kada je iduće godine uspjela njegovu fotografiju staviti na naslovnicu časopisa Forbes kao čovjeka koji je uspio povećati kapitalizaciju PerkinElmera s 1,5 milijardi dolara na 24 milijarde dolara 8 . (“Tony White je jadnog PerkinElmera pretvorio u visokotehnološkog hvatača gena.”) Tonyja su također progonile moje društvene aktivnosti.
Održao sam govor otprilike jednom tjedno, prihvaćajući mali dio od ogromnog broja poziva koje sam stalno primao jer je svijet želio znati za naš rad. Tony se čak požalio upravnom odboru PerkinElmera, koji je tada bio preimenovan u PE Corporation, da moja putovanja i pojavljivanja krše korporativna pravila. Tijekom dvotjednog odmora (o mom vlastitom trošku) u mom domu na Cape Codu, Tony je odletio u Celeru s financijskim direktorom Dennisom Wingerom i glavnim savjetnikom Applere Williamom Sauchom kako bi intervjuirali moje najbolje zaposlenike o "Venterovoj učinkovitosti upravljanja". Nadali su se da će skupiti dovoljno prljavštine da opravdaju moj otkaz. White je bio šokiran kad su svi rekli da će i oni odustati ako ja odustanem. To je izazvalo dosta napetosti unutar našeg tima, ali nas je i zbližilo više nego ikada. Svaku pobjedu smo bili spremni slaviti kao da nam je zadnja.
Nakon objave sekvence genoma muhe - do tada najveće sekvence u povijesti - Gene, Ham, Mark i ja nazdravili smo što smo izdržali Tonyja Whitea dovoljno dugo da naš uspjeh bude priznat. Dokazali smo da će naša metoda raditi i pri sekvenciranju ljudskog genoma. Čak i da Tony White sljedeći dan prestane financirati, znali smo da će naše glavno postignuće ostati s nama. Više od svega želio sam napustiti Celeru i ne morati imati posla s Tonyjem Whiteom, ali budući da sam želio još više sekvencirati genom Homo sapiensa, morao sam napraviti kompromis. Pokušao sam najbolje što sam mogao zadovoljiti Whitea, samo da nastavim posao i dovršim svoj plan.
Bilješke
1. Shreeve J. Rat genoma: Kako je Craig Venter pokušao uhvatiti šifru života i spasiti svijet (New York: Ballantine, 2005.), str. 285.
2. Ashburner M. Pobijedio za sve: Kako je sekvenciran genom Drosophile (Cold Spring Harbor Laboratory Press, 2006.), str. 45.
3. Shreeve J. Rat genoma, str. 300.
4. Ashburner M. Pobijedio za sve, str. 55.
5. Sulston J., Ferry G. Zajednička nit (London: Corgi, 2003.), str. 232.
6. Adams M. D., Celniker S. E. i sur. "The Genome Sequence of Drosophila Melanogaster", Science, br. 287, 2185–95, 24. ožujka 2000.
7. Gillis J. “Hoće li ovaj MAVERICK otključati najveće znanstveno otkriće svog doba? Kopernik, Newton, Einstein i VENTER?”, USA Weekend, 29. – 31. siječnja 1999.
8. Ross P. E. “Gene Machine”, Forbes, 21. veljače 2000.
Craig Venter