Le schéma d'obtention des hybrides de la première génération. La première loi de Mendel est la loi de domination. La loi de pureté des gamètes et sa justification cytologique
6. Qu'est-ce que l'ontogenèse. Quelles sont les périodes de l'ontogenèse ? Donnez une brève description du développement post-embryonnaire.
1. Niveaux des organisations fauniques (liste).
2. Nommez les organites cellulaires à deux membranes et non membranaires. Indiquez les caractéristiques de leur structure et de leur fonction.
3. Quels sont les avantages du processus de reproduction sexuée par rapport à l'asexuée.
4. Troisième loi de Mendel. Indiquez le ratio de la progéniture par génotype et phénotype.
5. Nommer les centres d'origine et de diversité des plantes cultivées selon Vavilov. Où est stockée la collection de graines de plantes poussant sur Terre, quelle est sa signification.
6. Qu'est-ce que l'ontogenèse. Quelles sont les périodes de l'ontogenèse ? Donnez une brève description du développement post-embryonnaire.
le corps de l'hôte. Mesures préventives.
2. Les forces motrices de l'évolution et leurs relations.
5. Déterminez le grossissement du microscope. Préparez-le pour le travail.
Billet numéro 6
1. Le système circulatoire des vertébrés.
2.1. Croisement monohybride.
2. Les première et deuxième lois de Mendel.
6.À l'aide de matériel d'herbier, regroupez les plantes en familles.
Billet numéro 7
1. Départements du système nerveux : central et périphérique. 2. L'habitat des organismes.
Biologie, au secours !!!1. Science qui étudie les modèles de transmission des traits par héritage
2. Les modèles génétiques ont d'abord été formulés
3. Chromosomes appariés, dont chacun va au corps de l'un des parents
4. Chromosomes contenant un ensemble de gènes identiques
5. Chromosomes situés dans le même organisme, mais différents en taille, forme et ensemble de gènes
6. Gènes situés dans les mêmes régions de chromosomes homologues, responsables de la synthèse des mêmes protéines, mais ayant une séquence nucléotidique différente
7. Quel sera le nom des gènes codant pour la même protéine (par exemple, l'hémoglobine ou la mélanine), mais contenant une séquence d'acides aminés différente
8. En croisement monohybride, les signes sont recherchés
9. Selon le gène de couleur (dans les expériences de Mendel), les pois avaient le nombre de traits
10. Zygote qui a reçu deux allèles différents
11. Zygote qui a reçu les deux mêmes allèles
12. Le gène indiqué lorsqu'il est traversé par une lettre minuscule
13. Quel est le nom de l'allèle, indiqué lors du croisement d'une lettre majuscule (majuscule)
14. Un allèle qui n'apparaît phénotypiquement qu'à l'état homozygote
15. Allèle, qui se manifeste à l'extérieur dans tous les cas, s'il est présent dans le génotype d'un individu
16. Génotype zygote désigné AA
17. Génotype zygote désigné Aa
18. Génotype du zygote, désigné aa
19. L'ensemble des signes extérieurs du corps
20. La totalité de toutes les caractéristiques héréditaires de l'organisme
21. L'ensemble de tous les gènes d'une espèce ou d'une population
22. Le fractionnement des hybrides de la deuxième génération suit la formule 3 : 1
23. Génotype des formes parentales de pois ("clean lines") dans les expériences de Mendel
24. Génotype des descendants (hybrides de la 1ère génération) chez les pois dans les expériences de Mendel
25. Couleur pois de première génération
26. F1 Mendel obtenu par pollinisation
27. F2 Mendel obtenu par pollinisation
28. Type de clivage lors du croisement hétérozygote et homozygote pour récessif
29. Formes monohétérozygotes ... variétés de gamètes
30. Hybrides avec croisement monohybride et forme de dominance complète ... classes phénotypiques
31. Les lois de Mendel traitaient de ce type d'héritage
32. Les gènes non alléliques situés sur le même chromosome, s'il n'y a pas de croisement, sont hérités
33. L'auteur de la théorie chromosomique de l'hérédité
34. Chromosomes de taille et de forme différentes chez les hommes et les femmes
35. Chromosomes qui ne diffèrent pas par leur taille et leur forme
36. Les gènes « liés au sexe » sont localisés
37. L'hémophilie et le daltonisme chez l'homme et l'hérédité de la couleur du pelage chez le chat en sont des exemples
38. Les gènes de ces traits sont localisés
39. Les mammifères mâles et la drosophile sont hétérozygotes pour les traits liés au sexe
40. Modifications accidentelles du matériel génétique d'un individu
41. Modification du nombre de chromosomes
42. Changement d'un ou plusieurs nucléotides
43. Modifications de la séquence d'acides aminés des protéines
44. L'auteur de la loi des séries homologues de variation héréditaire
45. La principale méthode de la génétique, inapplicable en génétique humaine
46. La méthode de la génétique humaine, basée sur la coloration des chromosomes et l'étude ultérieure de leur taille et de leur forme
47. La méthode de la génétique humaine, basée sur l'analyse des pedigrees et traçant la transmission d'un certain trait
48. La méthode de la génétique humaine, basée sur l'analyse des manifestations phénotypiques des traits chez des jumeaux identiques
49. La méthode de la génétique humaine, basée sur l'analyse de l'activité enzymatique des protéines qui catalysent d'importants processus physiologiques
50. Substances provoquant des mutations
Introduction.
La génétique est une science qui étudie les lois de l'hérédité et de la variabilité des organismes vivants.
L'homme a noté depuis longtemps trois phénomènes liés à l'hérédité : d'abord, la similitude des traits des descendants et des parents ; deuxièmement, les différences entre certains traits (parfois nombreux) de la progéniture et les traits parentaux correspondants ; troisièmement, l'apparition dans la descendance de caractères qui n'appartenaient qu'à des ancêtres lointains. La continuité des traits entre les générations est assurée par le processus de fécondation. Depuis des temps immémoriaux, l'homme a spontanément utilisé les propriétés de l'hérédité à des fins pratiques - pour la sélection de variétés de plantes cultivées et de races d'animaux domestiques.
Les premières idées sur le mécanisme de l'hérédité ont été exprimées par les anciens scientifiques grecs Démocrite, Hippocrate, Platon, Aristote. L'auteur de la première théorie scientifique de l'évolution J.-B. Lamarck a utilisé les idées des anciens scientifiques grecs pour expliquer ce qu'il a postulé au tournant des XVIIIe et XIXe siècles. le principe du transfert de nouveaux traits acquis au cours de la vie d'un individu à sa descendance. C. Darwin a avancé la théorie de la pangenèse, expliquant l'hérédité des caractères acquis
Charles Darwin défini hérédité comme propriété de tous les organismes vivants de transmettre leurs caractéristiques et propriétés de génération en génération, et variabilité comme la propriété de tous les organismes vivants d'acquérir de nouvelles caractéristiques au cours du processus de développement individuel.
Les traits sont hérités par la reproduction. Avec la reproduction sexuée, de nouvelles générations apparaissent à la suite de la fécondation. Les fondements matériels de l'hérédité sont contenus dans les cellules germinales. En reproduction asexuée ou végétative, une nouvelle génération se développe soit à partir de spores unicellulaires, soit à partir de formations multicellulaires. Et avec ces formes de reproduction, la connexion entre les générations s'effectue à travers les cellules, qui contiennent les fondements matériels de l'hérédité (unités élémentaires de l'hérédité) - les gènes - sont des sections de l'ADN des chromosomes.
L'ensemble des gènes qu'un organisme reçoit de ses parents constitue son génotype. L'ensemble des traits externes et internes est un phénotype. Le phénotype se développe à la suite de l'interaction du génotype et des conditions environnementales. D'une manière ou d'une autre, la base est constituée des traits porteurs de gènes.
Les modèles par lesquels les signes sont transmis de génération en génération ont été découverts pour la première fois par le grand scientifique tchèque Gregor Mendel. Il a découvert et formulé trois lois de l'hérédité qui ont formé la base de la génétique moderne.
La vie et la recherche scientifique de Gregor Johann Mendel.
Moine morave et phytogénéticien. Johann Mendel est né en 1822 dans la ville de Heinzendorf (aujourd'hui Gincice en République tchèque), où son père possédait une petite parcelle paysanne. Gregor Mendel, selon le témoignage de ceux qui l'ont connu, était en effet une personne aimable et agréable. Après avoir reçu son éducation élémentaire à l'école du village local et plus tard, après avoir obtenu son diplôme du Collège des Piaristes de Leipzig, il a été admis en 1834 au Troppown Imperial-Royal Gymnasium dans la première classe de grammaire. Quatre ans plus tard, les parents de Johann, à la suite d'une confluence de nombreux événements malheureux qui se sont rapidement succédé, ont été complètement privés de la possibilité de rembourser les dépenses nécessaires liées à l'éducation, et leur fils, n'ayant alors que 16 ans, a été contraint de s'occuper en toute indépendance de son propre entretien. ... En 1843, Mendel fut admis au monastère augustinien de Saint-Thomas à Altbrunn, où il prit le nom de Gregor. En 1846, Mendel a également assisté à des conférences sur l'agriculture, l'horticulture et la viticulture à l'Institut philosophique de Brunn. En 1848, après avoir terminé un cours de théologie, Mendel reçut, avec un profond respect, la permission d'étudier pour les examens de doctorat. Lorsque, l'année suivante, il renforça son intention d'être examiné, il reçut l'ordre de prendre la place du partisan du gymnase impérial-royal de Znaim, qu'il suivit avec joie.
En 1851, l'abbé du monastère envoya Mendel étudier à l'Université de Vienne, où il étudia entre autres la botanique. Après avoir obtenu son diplôme universitaire, Mendel a enseigné les sciences dans une école locale. Grâce à cette étape, sa situation financière a radicalement changé. Dans le bien-être bénéfique si nécessaire de l'existence physique pour chaque poursuite, le courage et la force lui revinrent avec une profonde révérence, et pendant l'année d'essai, il étudia les matières classiques prescrites avec beaucoup de diligence et d'amour. Pendant ses heures libres, il s'occupait d'une petite collection botanique et minéralogique, qui était à sa disposition dans le monastère. Sa passion pour le domaine des sciences naturelles devenait d'autant plus grande, plus il avait d'opportunités de s'abandonner à lui. Bien que celui mentionné dans ces leçons ait été privé de toute orientation, et que le chemin de l'autodidacte ici, comme dans aucune autre science, soit difficile et mène lentement au but, néanmoins pendant ce temps Mendel a acquis un tel amour pour l'étude de la nature que il n'a pas épargné ses efforts pour combler ses nouvelles lacunes en s'autodidacte et en suivant les conseils de personnes ayant une expérience pratique. Le 3 avril 1851, le « corps enseignant » de l'école décide d'inviter M. Gregor Mendel, chanoine du monastère de St. Les succès pomologiques de Gregor Mendel lui ont valu le droit à un titre stellaire et à un poste temporaire de partisan d'histoire naturelle dans la classe préparatoire de l'École technique. Au premier semestre de ses études, il n'a étudié que dix heures par semaine et uniquement avec Doppler. Au second semestre, il a étudié vingt heures par semaine. Parmi eux, dix - en physique avec Doppler, cinq par semaine - en zoologie avec Rudolf Kner. Onze heures par semaine - botanique avec le professeur Fenzl : en plus des cours de morphologie et de taxonomie, il a également suivi un atelier spécial sur la description et la définition des plantes. Au troisième semestre, il s'inscrit à trente-deux heures de cours par semaine : dix heures de physique à Doppler, dix heures de chimie à Rottenbacher : chimie générale, chimie médicinale, chimie pharmacologique, et un atelier de chimie analytique. Cinq pour la zoologie de Kner. Six heures de formation avec Unger, l'un des premiers cytologistes au monde. Dans ses laboratoires, il étudie l'anatomie et la physiologie végétales et suit un atelier sur les techniques de microscopie. Et encore - une fois par semaine au Département de Mathématiques - un atelier sur le logarithme et la trigonométrie.
1850, la vie allait bien. Mendel pouvait déjà subvenir à ses besoins et jouissait d'un grand respect de la part de ses collègues, car il s'acquittait bien de ses fonctions et était très agréable à qui parler. Ses disciples l'aimaient.
En 1851, Gregor Mendel se précipita sur la question cardinale de la biologie - le problème de la variabilité et de l'hérédité. C'est alors qu'il a commencé à mener des expériences sur la culture dirigée des plantes. Mendel a apporté diverses plantes de loin et près des environs de Brunn. Il a cultivé des plantes en groupes dans une partie du jardin du monastère spécialement désignée pour chacun d'eux dans diverses conditions extérieures. Il était engagé dans des observations météorologiques minutieuses. Gregor a mené la plupart de ses expériences et observations avec des pois, qu'il semait depuis 1854 chaque printemps chaque printemps dans un petit jardin sous les fenêtres de la prélature. Il s'est avéré facile d'établir une expérience d'hybridation claire sur les pois. Pour ce faire, il vous suffit d'ouvrir une grande fleur, mais pas encore mûre, avec une pince à épiler, de couper les anthères et de déterminer indépendamment la "paire" à croiser. L'autofécondation étant exclue, les variétés de pois sont, en règle générale, des "lignées propres" aux caractéristiques constantes qui ne changent pas de génération en génération, qui sont très clairement définies. Mendel a distingué les traits qui déterminent les différences intervariétales : la couleur de la peau des grains mûrs et, séparément, des grains immatures, la forme des pois mûrs, la couleur de la "protéine" (endosperme), la longueur de l'axe de la tige , l'emplacement et la couleur des bourgeons. Il a utilisé plus de trente variétés dans l'expérience, et chacune des variétés a été préalablement soumise à un test de deux ans pour la "constance", pour la "constance des traits", pour la "pureté du sang" - en 1854 et en 1855. Les expériences avec les pois ont duré huit ans. Des centaines de fois en huit floraisons de ses propres mains, il a soigneusement coupé les anthères et, tapant le pollen des étamines d'une fleur d'une autre variété sur une pince à épiler, l'a appliqué sur le stigmate du pistil. Dix mille passeports ont été délivrés pour dix mille plantes obtenues à la suite de croisements et d'hybrides autogames. Les entrées y sont soignées : quand la plante mère a été cultivée, quelles fleurs elle avait, dont le pollen a été fécondé, quels pois - jaunes ou verts, lisses ou ridés - ont été obtenus, quelles fleurs - la couleur des bords, la couleur en le centre - fleuri lorsque les graines ont été reçues combien d'entre elles sont jaunes, combien sont vertes, rondes, ridées, combien d'entre elles sont sélectionnées pour la plantation, quand elles sont plantées, etc.
Le résultat de ses recherches fut le rapport "Expériences sur les hybrides de plantes", qui fut lu par un naturaliste brunnien en 1865. Le rapport dit : « La raison de la mise en place des expériences auxquelles cet article est consacré était le croisement artificiel de plantes ornementales, qui a été réalisé afin d'obtenir de nouvelles formes, de couleurs différentes. Car mettre en place d'autres expériences afin de retracer le développement des hybrides dans leur progéniture a donné une impulsion à la régularité remarquable avec laquelle les formes hybrides retournaient constamment à leurs formes ancestrales. Comme cela arrive souvent dans l'histoire des sciences, l'œuvre de Mendel n'a pas été immédiatement reconnue par ses contemporains. Les résultats de ses expériences ont été publiés lors d'une réunion de la Société des sciences naturelles de la ville de Brunn, puis publiés dans le journal de cette société, mais les idées de Mendel à cette époque n'ont pas trouvé de soutien. Un numéro du magazine décrivant l'œuvre révolutionnaire de Mendel prend la poussière dans les bibliothèques depuis trente ans. Ce n'est qu'à la fin du XIXe siècle que des scientifiques traitant des problèmes d'hérédité découvrent les œuvres de Mendel, et il peut recevoir (déjà à titre posthume) la reconnaissance qu'il mérite.
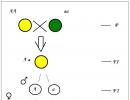
La première loi de Mendel
Le croisement de deux organismes s'appelle hybridation, la progéniture issue du croisement de deux individus d'hérédité différente est appelée hybride, et un individu - un hybride.Monohybride s'appelle le croisement de deux organismes qui diffèrent l'un de l'autre par une paire de traits alternatifs (mutuellement exclusifs). Par conséquent, avec un tel croisement, les modèles d'hérédité de seulement deux traits sont tracés, dont le développement est dû à une paire de gènes alléliques. Tous les autres signes caractéristiques de ces organismes ne sont pas pris en compte.
Si vous croisez des plants de pois avec des graines jaunes et vertes, tous les hybrides résultants auront des graines jaunes. La même image est observée lors du croisement de plantes avec une forme de graine lisse et ridée; tous les descendants de la première génération auront une forme de graine lisse :
Par conséquent, la première loi de Mendel a été nommée La loi d'uniformité des hybrides de première génération.
Si nous utilisons les termes apparus de nombreuses années après les travaux de Mendel, nous pouvons alors dire que les cellules des plants de pois d'une variété contiennent deux gènes de couleur jaune uniquement et les gènes de plantes d'une autre variété - deux gènes de couleur verte uniquement. Les gènes responsables du développement d'un caractère (par exemple, la couleur des graines) sont appelés gènes alléliques... Par conséquent, dans un hybride de la première génération, un seul de chaque paire de traits alternatifs se développe. Le deuxième signe semble disparaître, n'apparaît pas. Le phénomène de prédominance dans un hybride avecsigne d'un des parents G. Mendel nommé domrationnement. Un trait qui se manifeste dans un hybride de la première génération et supprime le développement d'un autre trait a été nommé dominant, et le contraire, c'est-à-dire supprimé, signe - récessif. Gregor Mendel a décidé de remplacer la description des caractères végétaux par le code abstrait "A, B, C, D, E, F, G" et "a, b, c, d, e, f, g" puis d'observer les sort d'une paire de traits, il est passé à regarder deux, trois, quatre paires en même temps. Grand A, B, C, D, E, F, G, il marqua les traits dominants ; petit a, b, c, d, e, f, g - récessif. S'il y a deux gènes alléliques identiques dans le génotype d'un organisme (zygote), les deux sont dominants ou les deux sont récessifs (AA ou aa), un tel organisme est appelé homozygote. Si sur une paire de gènes alléliques, l'un est dominant et l'autre récessif (Aa), alors un tel organisme est appelé hétérozygote.
La loi de division, ou la deuxième loi de Mendel.
Si les descendants de la première génération, identiques dans le trait étudié, sont croisés entre eux, alors dans la deuxième génération les traits des deux parents apparaissent dans un certain rapport numérique : 3/4 des individus auront un trait dominant, ¼ récessif:

Le phénomène dans lequel le croisement des hétérozygotesindividus conduit à la formation de descendants, dont certainsrogo porte une caractéristique dominante, et une partie- récessifny, est appelé fractionnement. Par conséquent, le trait récessif chez les hybrides de la première génération n'a pas disparu, mais a seulement été supprimé et se manifestera dans la deuxième génération d'hybrides.
Hypothèse de pureté des gamètes. Mendel a suggéré que lors de la formation des hybrides, les facteurs héréditaires ne se mélangent pas, mais restent inchangés. Dans l'hybride, les deux facteurs sont présents - dominant et récessif, mais le facteur héréditaire dominant se manifeste sous la forme d'un trait, tandis que le facteur récessif est supprimé. La communication entre les générations au cours de la reproduction sexuée s'effectue à travers les cellules sexuelles - Haméta. Par conséquent, il faut supposer que chaque gamète ne porte qu'un seul facteur d'une paire. Ensuite, lors de la fécondation, la fusion de deux gamètes, chacun porteur d'un facteur héréditaire récessif, conduira à la formation d'un organisme doté d'un trait récessif qui se manifeste phénotypiquement. La fusion de gamètes, chacun porteur d'un facteur dominant, ou de deux gamètes dont l'un contient un facteur dominant et l'autre un facteur récessif, conduira au développement d'un organisme doté d'un trait dominant.
Mendel a expliqué la division de la progéniture lors du croisement d'individus hétérozygotes par le fait que les gamètes sont génétiquement purs, c'est-à-dire qu'ils ne portent qu'un seul gène d'une paire allélique. Hypothèse(maintenant appelé la loi) puretégamètes peut être formulé comme suit : lors de la formation des cellules germinales, un seul gène d'une paire allélique pénètre dans chaque gamète.
Pourquoi et comment cela se produit-il ? On sait que chaque cellule du corps possède exactement le même ensemble diploïde de chromosomes. Deux chromosomes homologues contiennent deux gènes identiques. Les gamètes génétiquement "purs" se forment comme suit : lorsque les gamètes mâles et femelles fusionnent, un hybride avec un ensemble diploïde (double) de chromosomes est obtenu.
Comme on peut le voir sur le schéma (annexe 2), le zygote reçoit la moitié des chromosomes de l'organisme paternel et l'autre moitié du maternel.
Au cours du processus de formation des gamètes chez un hybride, des chromosomes homologues au cours de la première division méiotique pénètrent également dans différentes cellules.
Pour cette paire allélique, deux variétés de gamètes sont formées. Au cours de la fécondation, des gamètes portant le même allèle ou des allèles différents se rencontrent accidentellement. En raison de la probabilité statistique, avec un nombre suffisamment important de gamètes dans la descendance, 25 % des génotypes seront homozygotes dominants, 50 % - hétérozygotes, 25 % - homozygotes récessifs, c'est-à-dire le rapport 1AA : 2Aa : 1aa.
Ainsi, selon le phénotype, la descendance de la deuxième génération lors du croisement monohybride est répartie dans un rapport de 3 : 1 (¾ individus avec un trait dominant, ¼ individus avec un trait récessif).
La base cytologique du clivage des caractères dans le croisement monohybride est la divergence des chromosomes homologues aux différents pôles de la cellule et la formation de cellules germinales haploïdes lors de la méiose.
Dans les exemples discutés ci-dessus, la règle d'uniformité a été exprimée dans le fait que tous les hybrides étaient extérieurement similaires à l'un des parents. Ce n'est pas toujours le cas. Souvent, les signes dans les formes hétérozygotes sont de nature intermédiaire, c'est-à-dire la domination peut ne pas être complète. Schéma de croisement de deux formes héréditaires de la plante Night Beauty :


L'un d'eux a des fleurs rouges (et c'est la caractéristique dominante), et l'autre est blanc. Le diagramme montre que tous les hybrides de première génération ont des fleurs roses. Dans la deuxième génération, la division se produit dans un rapport de 1: 2: 1, c'est-à-dire une fleur rouge (homozygote), deux fleurs roses (hétérozygote), une blanche (homozygote). Ce phénomène est appelé dominance incomplète.
En cas de dominance incomplète, le gène dominant à l'état hétérozygote ne supprime pas toujours complètement le gène récessif. Dans certains cas, un hybride Fi ne reproduit entièrement aucun des traits parentaux et le trait est de nature intermédiaire avec plus ou moins de déviation vers un état dominant ou récessif. Mais tous les individus de cette génération sont uniformes dans cette caractéristique. La domination incomplète est répandue. Il a été trouvé dans l'étude de l'hérédité de la couleur des fleurs chez les mufliers, de la couleur de la laine chez les bovins et les moutons, les traits biochimiques chez l'homme, etc. Les traits intermédiaires résultant d'une dominance incomplète représentent souvent une valeur esthétique ou matérielle pour l'homme. La question se pose: est-il possible de reproduire par sélection, par exemple, une variété de beauté nocturne avec une couleur de fleurs rose? Evidemment non, car ce trait ne se développe que chez les hétérozygotes et lorsqu'ils sont croisés entre eux, il se produit toujours un clivage :
La domination incomplète est répandue. Il a été trouvé dans l'étude de l'hérédité de la couleur des fleurs chez les mufliers, de la couleur de la laine chez les bovins et les moutons, les traits biochimiques chez l'homme, etc. Les traits intermédiaires résultant d'une dominance incomplète représentent souvent une valeur esthétique ou matérielle pour l'homme. La question se pose: est-il possible de reproduire par sélection, par exemple, une variété de beauté nocturne avec une couleur de fleurs rose? Evidemment non, car ce trait ne se développe que chez les hétérozygotes et lorsqu'ils sont croisés, une scission se produit toujours.
La loi de combinaison indépendante, ou la troisièmeLa loi de Mendel.
L'étude de Mendel sur l'hérédité d'une paire d'allèles a permis d'établir un certain nombre de schémas génétiques importants : le phénomène de dominance, l'immuabilité des allèles récessifs chez les hybrides, la division de la progéniture des hybrides dans un rapport de 3: 1, et aussi de supposer que les gamètes sont génétiquement purs, c'est-à-dire qu'ils ne contiennent qu'un seul gène de paires alléliques. Cependant, les organismes diffèrent par de nombreux gènes. Il est possible d'établir les modèles d'hérédité de deux paires de traits alternatifs et plus en dihybride ou croisement polyhybride, c'est-à-dire croisement de formes parentales différant par deux paires de traits.
Pour le croisement dihybride, Mendel a pris des plants de pois homozygotes qui diffèrent par deux indicateurs - la couleur des graines (jaune, verte) et la forme des graines (lisse, ridée). Caractéristiques dominantes - coloration jaune (UNE) et forme lisse (V) des graines. Chaque plante forme une variété de gamètes pour les allèles étudiés : Lorsque les gamètes fusionnent, tous les descendants seront uniformes :


|
Treillis de pannet |
||||||

Il est facile de calculer que selon le phénotype, la progéniture est divisée en 4 groupes : 9 jaunes lisses, 3 jaunes ridées, 3 vertes lisses, 1 jaune ridée (9 : 3 : 3 : 1). Si nous prenons en compte les résultats du fractionnement pour chaque paire de caractères séparément, il s'avère que le rapport du nombre de graines jaunes au nombre de graines vertes et le rapport des graines lisses aux graines ridées pour chaque paire est de 3: 1 . Cela peut être exprimé algébriquement comme le carré du binôme
(3 + 1) ² = 3² + 2 · 3 + 1² ou 9 + 3 + 3 + 1
Ainsi, dans un croisement dihybride, chaque paire de traits lors de la division de la descendance se comporte de la même manière que dans un croisement monohybride, c'est-à-dire indépendamment de l'autre paire de traits.
Lors de la fécondation, les gamètes sont combinés selon les règles des combinaisons aléatoires, mais avec une probabilité égale pour chacun. Dans les zygotes résultants, diverses combinaisons de gènes apparaissent. Nous pouvons maintenant formuler la troisième loi de Mendel : lors du croisement de deux individus homozygotes, différentsséparés les uns des autres par deux ou plusieurs paires de caractères, gènes etleurs traits correspondants sont hérités indépendammentles uns des autres et se combinent de toutes les manières possiblescombinaisons.
Les lois de Mendel servent de base à l'analyse du clivage dans des cas plus complexes : lorsque les individus diffèrent par trois, quatre paires de caractères ou plus.
Conditions de respect des lois successorales de Mendel
Les lois découvertes par Gregor Mendel ne sont pas toujours applicables en génétique. Il existe de nombreuses conditions pour se conformer aux lois de Mendel. Pour de tels cas, il existe d'autres lois (par exemple : la loi de Morgan), ou des explications.
Formulons les conditions de base pour l'observation des lois de l'hérédité.
Pour respecter la loi d'homogénéité des hybrides de première génération, il faut que :
les organismes parentaux étaient homozygotes;
les gènes de différents allèles étaient situés dans différents chromosomes, et non dans un seul (sinon, le phénomène de « hérédité liée » peut se produire).
La loi de fractionnement sera respectée si
chez les hybrides, les facteurs héréditaires restent inchangés ;
La loi de distribution indépendante des gènes dans la descendance et l'émergence de diverses combinaisons de ces gènes lors du croisement dihybride n'est possible qu'en cas.
si des paires de gènes alléliques sont situées dans différentes paires de chromosomes homologues.
La violation de ces conditions peut conduire soit à l'absence de clivage en deuxième génération, soit à un clivage en première génération ; ou à la distorsion du rapport des différents génotypes et phénotypes. Les lois de Mendel sont universelles pour tous les organismes diploïdes se reproduisant sexuellement. En général, ils sont valables pour des gènes autosomiques avec une pénétrance complète (100% de fréquence de manifestation du trait analysé ; 100% de pénétrance implique que le trait s'exprime chez tous les porteurs de l'allèle qui détermine le développement de ce trait) et une expressivité constante ; une expressivité constante implique que la sévérité phénotypique d'un trait est la même ou approximativement la même chez tous les porteurs de l'allèle qui détermine le développement de ce trait.
Conclusion.
Les lois de Gregor Mendel sont actuellement largement utilisées dans la sélection de plantes, d'animaux et de micro-organismes, en médecine, en génie génétique et dans de nombreuses autres branches de la vie humaine, ainsi que pour résoudre des problèmes de génétique.
Il est important de noter que Mendel a formulé des lois et tiré des conclusions à une époque où l'on ne savait rien de l'ADN, des gènes et des chromosomes. Cependant, il s'est avéré avoir tout à fait raison, et bien que pas immédiatement, ses théories ont été reconnues et prises comme base du développement de la science de la génétique.
Théorie mendélienne de l'hérédité, c'est-à-dire l'ensemble des idées sur les déterminants héréditaires et la nature de leur transmission des parents aux descendants, dans son sens, est directement opposé aux théories domaineléennes, en particulier, la théorie de la pangenèse proposée par Darwin. Conformément à cette théorie, les signes des parents sont directs, c'est-à-dire de toutes les parties du corps, transmis à la progéniture. Par conséquent, le caractère du trait du descendant devrait dépendre directement des propriétés du parent. Ceci contredit complètement les conclusions de Mendel : les déterminants de l'hérédité, c'est-à-dire les gènes sont présents dans le corps relativement indépendamment de lui-même. La nature des traits (phénotype) est déterminée par leur combinaison aléatoire. Ils ne sont modifiés par aucune partie du corps et sont dans une relation de dominance récessive. Ainsi, la théorie mendélienne de l'hérédité s'oppose à l'idée d'héritage des caractères acquis au cours du développement individuel.
Les expériences de Mendel ont servi de base au développement de la génétique moderne - une science qui étudie deux propriétés principales d'un organisme - l'hérédité et la variabilité. Il a réussi à identifier les modèles d'héritage grâce à des approches méthodologiques fondamentalement nouvelles :
1) Mendel a choisi avec succès l'objet de la recherche ;
2) il a analysé l'héritage de traits individuels dans la progéniture de plantes croisées, différant par une, deux et trois paires de traits alternatifs contrastés. A chaque génération, un enregistrement a été tenu séparément pour chaque paire de ces caractéristiques ;
3) il a non seulement enregistré les résultats obtenus, mais a également effectué leur traitement mathématique.
Les méthodes de recherche simples énumérées constituaient une méthode hybride fondamentalement nouvelle pour l'étude de l'hérédité, qui est devenue la base de nouvelles recherches en génétique.
Bibliographie.
Biologie générale : Manuel pour les 9e à 10e années. mer école / Polyansky Yu.I., Brown A.D., Verzilin N.M. et autres; M. : Education, 1987.-287 p. : ill.
Mendel est arrivé à la conclusion que la prédisposition à l'accouchement ... affecte les fréquences des génotypes dans la population. Les lois Mendel ne nous dit rien sur les fréquences...La première loi de Mendel
Le croisement de deux organismes s'appelle hybridation, la progéniture issue du croisement de deux individus d'hérédité différente est appelée hybride, et un individu - un hybride. Mono hybride s'appelle le croisement de deux organismes qui diffèrent l'un de l'autre par une paire de traits alternatifs (mutuellement exclusifs). Par conséquent, avec un tel croisement, les modèles d'hérédité de seulement deux traits sont tracés, dont le développement est dû à une paire de gènes alléliques. Tous les autres signes caractéristiques de ces organismes ne sont pas pris en compte.
Si vous croisez des plants de pois avec des graines jaunes et vertes, tous les hybrides résultants auront des graines jaunes. La même image est observée lors du croisement de plantes avec une forme de graine lisse et ridée; tous les descendants de la première génération auront une forme de graine lisse :
Par conséquent, la première loi de Mendel a été nommée La loi d'uniformité des hybrides de première génération.
Si nous utilisons les termes apparus de nombreuses années après les travaux de Mendel, nous pouvons alors dire que les cellules des plants de pois d'une variété contiennent deux gènes de couleur jaune uniquement et les gènes de plantes d'une autre variété - deux gènes de couleur verte uniquement. Les gènes responsables du développement d'un caractère (par exemple, la couleur des graines) sont appelés gènes alléliques... Par conséquent, dans un hybride de la première génération, un seul de chaque paire de traits alternatifs se développe. Le deuxième signe semble disparaître, n'apparaît pas. Le phénomène de prédominance dans un hybride avec signe d'un des parents G. Mendel nommé dom rationnement. Un trait qui se manifeste dans un hybride de la première génération et supprime le développement d'un autre trait a été nommé dominant, et le contraire, c'est-à-dire supprimé, signe - récessif. Gregor Mendel a décidé de remplacer la description des caractères végétaux par le code abstrait "A, B, C, D, E, F, G" et "a, b, c, d, e, f, g" puis d'observer les sort d'une paire de traits, il est passé à regarder deux, trois, quatre paires en même temps. Grand A, B, C, D, E, F, G, il marqua les traits dominants ; petit a, b, c, d, e, f, g - récessif. S'il y a deux gènes alléliques identiques dans le génotype d'un organisme (zygote), les deux sont dominants ou les deux sont récessifs (AA ou aa), un tel organisme est appelé homozygote. Si sur une paire de gènes alléliques, l'un est dominant et l'autre récessif (Aa), alors un tel organisme est appelé hétérozygote.
La loi de division, ou la deuxième loi de Mendel.
Si les descendants de la première génération, identiques dans le trait étudié, sont croisés entre eux, alors dans la deuxième génération les traits des deux parents apparaissent dans un certain rapport numérique : 3/4 des individus auront un trait dominant, ¼ récessif:

Le phénomène dans lequel le croisement des hétérozygotes individus conduit à la formation de descendants, dont certains rogo porte une caractéristique dominante, et une partie - récessif ny, est appelé fractionnement. Par conséquent, le trait récessif chez les hybrides de la première génération n'a pas disparu, mais a seulement été supprimé et se manifestera dans la deuxième génération d'hybrides.
Hypothèse de pureté des gamètes. Mendel a suggéré que lors de la formation des hybrides, les facteurs héréditaires ne se mélangent pas, mais restent inchangés. Dans l'hybride, les deux facteurs sont présents - dominant et récessif, mais le facteur héréditaire dominant se manifeste sous la forme d'un trait, tandis que le facteur récessif est supprimé. La communication entre les générations au cours de la reproduction sexuée s'effectue à travers les cellules sexuelles - Ha méta. Par conséquent, il faut supposer que chaque gamète ne porte qu'un seul facteur d'une paire. Ensuite, lors de la fécondation, la fusion de deux gamètes, chacun porteur d'un facteur héréditaire récessif, conduira à la formation d'un organisme doté d'un trait récessif qui se manifeste phénotypiquement. La fusion de gamètes, chacun porteur d'un facteur dominant, ou de deux gamètes dont l'un contient un facteur dominant et l'autre un facteur récessif, conduira au développement d'un organisme doté d'un trait dominant.
Mendel a expliqué la division de la progéniture lors du croisement d'individus hétérozygotes par le fait que les gamètes sont génétiquement purs, c'est-à-dire qu'ils ne portent qu'un seul gène d'une paire allélique. Hypothèse(maintenant appelé la loi) pureté gamètes peut être formulé comme suit : lors de la formation des cellules germinales, un seul gène d'une paire allélique pénètre dans chaque gamète.
Pourquoi et comment cela se produit-il ? On sait que chaque cellule du corps possède exactement le même ensemble diploïde de chromosomes. Deux chromosomes homologues contiennent deux gènes identiques. Les gamètes génétiquement "purs" se forment comme suit : lorsque les gamètes mâles et femelles fusionnent, un hybride avec un ensemble diploïde (double) de chromosomes est obtenu.
Comme on peut le voir sur le schéma (annexe 2), le zygote reçoit la moitié des chromosomes de l'organisme paternel et l'autre moitié du maternel.
Au cours du processus de formation des gamètes chez un hybride, des chromosomes homologues au cours de la première division méiotique pénètrent également dans différentes cellules.
Pour cette paire allélique, deux variétés de gamètes sont formées. Au cours de la fécondation, des gamètes portant le même allèle ou des allèles différents se rencontrent accidentellement. En raison de la probabilité statistique, avec un nombre suffisamment important de gamètes dans la descendance, 25 % des génotypes seront homozygotes dominants, 50 % - hétérozygotes, 25 % - homozygotes récessifs, c'est-à-dire le rapport 1AA : 2Aa : 1aa.
Ainsi, selon le phénotype, la descendance de la deuxième génération lors du croisement monohybride est répartie dans un rapport de 3 : 1 (¾ individus avec un trait dominant, ¼ individus avec un trait récessif).
La base cytologique du clivage des caractères dans le croisement monohybride est la divergence des chromosomes homologues aux différents pôles de la cellule et la formation de cellules germinales haploïdes lors de la méiose.
Dans les exemples discutés ci-dessus, la règle d'uniformité a été exprimée dans le fait que tous les hybrides étaient extérieurement similaires à l'un des parents. Ce n'est pas toujours le cas. Souvent, les signes dans les formes hétérozygotes sont de nature intermédiaire, c'est-à-dire la domination peut ne pas être complète. Schéma de croisement de deux formes héréditaires de la plante Night Beauty :


L'un d'eux a des fleurs rouges (et c'est la caractéristique dominante), et l'autre est blanc. Le diagramme montre que tous les hybrides de première génération ont des fleurs roses. Dans la deuxième génération, la division se produit dans un rapport de 1: 2: 1, c'est-à-dire une fleur rouge (homozygote), deux fleurs roses (hétérozygote), une blanche (homozygote). Ce phénomène est appelé dominance incomplète.
En cas de dominance incomplète, le gène dominant à l'état hétérozygote ne supprime pas toujours complètement le gène récessif. Dans certains cas, un hybride Fi ne reproduit entièrement aucun des traits parentaux et le trait est de nature intermédiaire avec plus ou moins de déviation vers un état dominant ou récessif. Mais tous les individus de cette génération sont uniformes dans cette caractéristique. La domination incomplète est répandue. Il a été trouvé dans l'étude de l'hérédité de la couleur des fleurs chez les mufliers, de la couleur de la laine chez les bovins et les moutons, les traits biochimiques chez l'homme, etc. Les traits intermédiaires résultant d'une dominance incomplète représentent souvent une valeur esthétique ou matérielle pour l'homme. La question se pose: est-il possible de reproduire par sélection, par exemple, une variété de beauté nocturne avec une couleur de fleurs rose? Evidemment non, car ce trait ne se développe que chez les hétérozygotes et lorsqu'ils sont croisés entre eux, il se produit toujours un clivage :
La domination incomplète est répandue. Il a été trouvé dans l'étude de l'hérédité de la couleur des fleurs chez les mufliers, de la couleur de la laine chez les bovins et les moutons, les traits biochimiques chez l'homme, etc. Les traits intermédiaires résultant d'une dominance incomplète représentent souvent une valeur esthétique ou matérielle pour l'homme. La question se pose: est-il possible de reproduire par sélection, par exemple, une variété de beauté nocturne avec une couleur de fleurs rose? Evidemment non, car ce trait ne se développe que chez les hétérozygotes et lorsqu'ils sont croisés, une scission se produit toujours.
La loi de combinaison indépendante, ou la troisième La loi de Mendel.
L'étude de Mendel sur l'hérédité d'une paire d'allèles a permis d'établir un certain nombre de schémas génétiques importants : le phénomène de dominance, l'immutabilité des allèles récessifs chez les hybrides, la division de la progéniture des hybrides dans un rapport de 3: 1, et aussi de supposer que les gamètes sont génétiquement purs, c'est-à-dire qu'ils ne contiennent qu'un seul gène de paires alléliques. Cependant, les organismes diffèrent par de nombreux gènes. Il est possible d'établir les modèles d'hérédité de deux paires de traits alternatifs et plus en dihybride ou croisement polyhybride, c'est-à-dire croisement de formes parentales différant par deux paires de traits.
Un scientifique de la République tchèque Gregor Mendel (1822-1884), au cours de ses recherches sur le croisement de différentes variétés de pois, a révélé certains modèles qui sont maintenant largement connus sous le nom des trois lois de Mendel.
Première loi
Si des spécimens végétaux ont été obtenus dans la première génération en croisant des variétés parentales homozygotes (lignées pures) avec des traits différents, alors la progéniture est la même en génotype et en phénotype. Dans ce cas, un trait est compris comme toute qualité sélectionnée de plantes, par laquelle deux variétés peuvent être distinguées. La première loi de Mendel est également connue sous le nom de règle de dominance ou loi d'uniformité pour les hybrides de première génération. Le trait qui a été identifié dans la première génération a été appelé dominant, et la qualité qui a été supprimée et ne s'est pas manifestée était récessive.
Deuxième loi
Droit de division. Son essence réside dans le fait que lors du croisement, par exemple, lors d'une autofécondation, du même type d'hybrides de la première génération à la génération suivante, la progéniture est divisée selon le trait choisi (phénotype) dans un rapport de 3 : 1 en cas de dominance complète et 1: 2: 1 en cas de dominance incomplète.
La troisième loi
La troisième loi de Mendel traite de l'hérédité indépendante des traits, ou de la combinaison indépendante des gènes végétaux. S'il y a un croisement de spécimens végétaux qui diffèrent les uns des autres par deux ou plusieurs paires de caractères alternatifs, le matériel génétique et les caractères phénotypiques qu'ils déterminent sont hérités indépendamment les uns des autres et peuvent être combinés dans n'importe quelle combinaison.
Les expériences du scientifique tchèque sont devenues la base du développement de la génétique moderne. G. Mendel a pu découvrir les schémas de base de l'hérédité à l'aide des dernières approches méthodologiques, qui sont encore utilisées par tous les généticiens.
Dans ses expériences, tous les spécimens croisés appartenaient à la même espèce végétale. Il est important qu'en même temps, ils diffèrent clairement les uns des autres par une, deux, plusieurs paires de signes alternatifs (opposés). Ces traits étaient constants, c'est-à-dire qu'ils se transmettaient de génération en génération lors de croisements au sein d'une même espèce. Il était nécessaire d'étudier la progéniture de chaque paire d'hybrides individuellement et de faire un compte quantitatif des hybrides qui diffèrent par certaines paires de caractères alternatifs.
Ces méthodes de recherche ont constitué la base d'une nouvelle méthode hybridologique, avec l'avènement de laquelle a commencé l'ère de l'étude de l'hérédité et de la variabilité.
Les modèles d'hérédité des traits au cours de la reproduction sexuée ont été établis par G. Mendel. Il est nécessaire de bien comprendre le génotype et le phénotype, les allèles, l'homo- et l'hétérozygotie, la dominance et ses types, les types de croisements, et aussi d'établir des schémas.
Monohybride croisement est appelé, dans lequel les formes parentales diffèrent les unes des autres par une paire de caractères alternatifs contrastés.
Par conséquent, avec un tel croisement, les modèles d'hérédité de seulement deux variantes du trait sont tracés, dont le développement est dû à une paire de gènes alléliques. Des exemples de croisements monohybrides réalisés par G. Mendel sont des croisements de pois avec des traits alternatifs aussi clairement visibles que des fleurs violettes et blanches, la couleur jaune et verte des fruits immatures (haricots), la surface lisse et ridée des graines, la couleur jaune et verte, etc. .
Uniformité des hybrides de première génération (première loi de Mendel) En croisant des pois avec des fleurs violettes (AA) et blanches (aa), Mendel a découvert que toutes les plantes hybrides de première génération (F1) avaient des fleurs violettes (Figure 2).
Figure 2 – Schéma de croisement monohybride
Dans le même temps, la couleur blanche de la fleur n'apparaissait pas. En croisant des plantes qui ont une forme de graine lisse et ridée, les hybrides auront des graines lisses. G. Mendel a également découvert que tous les hybrides F 1 étaient uniformes (homogènes) pour chacun des sept caractères qu'il a étudiés. Par conséquent, chez les hybrides de la première génération, sur une paire de traits parentaux alternatifs, un seul apparaît, et le trait de l'autre parent semble disparaître.
Les signes alternatifs sont des signes opposés qui s'excluent mutuellement.
Mendel a qualifié la prédominance des traits de l'un des parents des hybrides F1 de dominance et le trait correspondant de dominant. Il a qualifié de récessifs les signes qui ne se manifestent pas chez les hybrides F1. Étant donné que tous les hybrides de première génération sont uniformes, ce phénomène a été appelé les premières lois de Mendel, ou la loi d'uniformité des hybrides de première génération, ainsi que la règle de dominance.
Il peut être formulé comme suit : lors du croisement de deux organismes appartenant à des lignées pures différentes (deux organismes homozygotes), différant l'un de l'autre par une paire de caractères alternatifs, toute la première génération d'hybrides sera uniforme et portera le caractère de l'un des les parents.
Chaque gène a deux états - "A" et "a", ils forment donc une paire, et chaque membre de la paire est appelé un allèle. Les gènes situés dans les mêmes loci (sections) de chromosomes homologues et déterminant le développement alternatif du même trait sont appelés alléliques.
Par exemple, la couleur pourpre et blanche d'une fleur de pois est dominante et récessive, respectivement, pour deux allèles (A et a) d'un gène. Du fait de la présence de deux allèles, deux états de l'organisme sont possibles : homo- et hétérozygote. Si un organisme contient les mêmes allèles d'un gène particulier (AA ou aa), alors il est dit homozygote pour ce gène (ou trait), et s'il est différent (Aa), alors hétérozygote. Par conséquent, un allèle est une forme de l'existence d'un gène. Un exemple de gène tri-allélique est un gène qui détermine le système de groupe sanguin ABO chez l'homme. Il y a plus d'allèles : pour le gène qui contrôle la synthèse de l'hémoglobine humaine, il y en a des dizaines.
À partir des graines de pois hybrides, Mendel a fait pousser des plantes, qu'il a soumises à l'autopollinisation, et les graines résultantes ont été semées à nouveau. Le résultat a été une deuxième génération d'hybrides, ou hybrides F2. Parmi ces derniers, un clivage a été trouvé pour chaque paire de caractères alternatifs dans un rapport d'environ 3 : 1, c'est-à-dire que les trois quarts des plantes avaient des caractères dominants (fleurs violettes, graines jaunes, graines lisses, etc.) et un quart étaient récessif (fleurs blanches, graines vertes, graines ridées, etc.). Par conséquent, le caractère récessif de l'hybride F1 n'a pas disparu, mais n'a été supprimé et n'a réapparu qu'à la deuxième génération. Cette généralisation a été appelée plus tard la deuxième loi de Mendel, ou la loi de la division.
Le clivage est un phénomène dans lequel le croisement d'individus hétérozygotes conduit à la formation d'une descendance, dont certaines sont dominantes et d'autres récessives.
Deuxième loi de Mendel : lorsque deux descendants de la première génération sont croisés (deux individus hétérozygotes) dans la deuxième génération, le clivage est observé dans un certain rapport numérique : selon le phénotype 3 : 1, selon le génotype 1 : 2 : 1 (Fig. 3).

Figure 3 - Schéma de séparation des panneaux
lors du croisement d'hybrides F 1
Le dédoublement des traits dans la descendance lors du croisement d'individus hétérozygotes a été expliqué par G. Mendel par le fait que les gamètes sont génétiquement purs, c'est-à-dire qu'ils ne portent qu'un seul gène d'une paire allélique. La loi de pureté des gamètes peut être formulée comme suit : lors de la formation des cellules germinales, un seul gène d'une paire allélique pénètre dans chaque gamète.
Il convient de garder à l'esprit que l'utilisation de la méthode hybridologique pour analyser l'hérédité des caractères chez toute espèce animale ou végétale implique les croisements suivants :
croisement de formes parentales (P), différant par un (croisement monohybride) ou plusieurs paires (croisement polyhybride) de caractères alternatifs et obtenant des hybrides de la première génération (F 1 );
croisement d'hybrides F 1 entre eux et obtention d'hybrides de deuxième génération (F 2 ) ;
analyse mathématique des résultats de croisement.
Par la suite, Mendel s'est tourné vers l'étude du croisement dihybride.
Traversée dihybride Est un croisement dans lequel deux paires d'allèles sont impliquées (les gènes appariés sont alléliques et ne sont localisés que dans les chromosomes homologues).
Au cours du croisement dihybride, G. Mendel a étudié l'hérédité des traits, dont les gènes qui se trouvent dans différentes paires de chromosomes homologues sont responsables. À cet égard, chaque gamète doit contenir un gène de chaque paire allélique.
Les hybrides hétérozygotes pour deux gènes sont appelés dihétérozygotes, et s'ils diffèrent par trois ou plusieurs gènes, ils sont respectivement tri- et polyhétérozygotes.
Des schémas plus complexes de croisements dihybrides, l'enregistrement des génotypes et des phénotypes de F 2 est réalisé à l'aide du réseau de Pennett. Prenons un exemple d'une telle croix. Pour le croisement, deux formes parentales homozygotes initiales ont été prises : la première forme avait des graines jaunes et lisses ; la seconde forme avait des graines vertes et ridées (Fig. 4).

Figure 4 - Croisement dihybride de plants de pois,
différentes dans la forme et la couleur des graines
La couleur jaune et les graines lisses sont des caractéristiques dominantes; la couleur verte et les graines ridées sont des symptômes récessifs. Hybrides de première génération - croisés les uns avec les autres. Dans la deuxième génération, une division phénotypique a été observée dans un rapport de 9 : 3 : 3 : 1, soit (3+1) 2, après autofécondation des hybrides F1 conformément à la loi de division, des graines ridées et vertes sont réapparues.
Les plantes mères dans ce cas ont les génotypes AABB et aabb, et le génotype des hybrides F 1 - AaBb, c'est-à-dire est dihétérozygote.
Ainsi, lors du croisement d'individus hétérozygotes différant par plusieurs paires de traits alternatifs, un clivage phénotypique est observé dans la descendance dans le rapport (3 + 1) n, où n est le nombre de paires de traits alternatifs.
Les gènes qui déterminent le développement de différentes paires de traits sont appelés non alléliques.
Les résultats des croisements dihybrides et polyhybrides dépendent du fait que les gènes qui déterminent les traits considérés sont situés dans le même ou dans des chromosomes différents. Mendel a découvert des traits dont les gènes se trouvaient dans différentes paires de chromosomes de pois homologues.
Dans la méiose, les chromosomes homologues de différentes paires sont combinés au hasard dans les gamètes. Si le chromosome paternel de la première paire pénètre dans le gamète, alors avec une probabilité égale le chromosome paternel et le chromosome maternel de la deuxième paire peuvent pénétrer dans ce gamète. Par conséquent, les traits dont les gènes se trouvent dans différentes paires de chromosomes homologues sont combinés indépendamment les uns des autres. Par la suite, il s'est avéré que sur les sept paires de caractères étudiées par Mendel chez le pois, dans lesquelles le nombre diploïde de chromosomes est de 2 n = 14, les gènes responsables de l'une des paires de caractères se trouvaient sur le même chromosome. Cependant, Mendel n'a pas trouvé de violation de la loi de l'hérédité indépendante, car le lien entre ces gènes n'a pas été observé en raison de la grande distance qui les sépare).
Sur la base de ses recherches, Mendel a déduit une troisième loi - la loi de l'hérédité indépendante des traits, ou combinaison indépendante des gènes.
Chaque paire de gènes alléliques (et les traits alternatifs qu'ils contrôlent) est héritée indépendamment les uns des autres.
La loi de combinaison indépendante des gènes est à la base de la variabilité combinatoire observée lors du croisement chez tous les organismes vivants. On note également que, contrairement à la première loi de Mendel, qui est toujours vraie, la seconde loi n'est valable que pour des gènes localisés dans différentes paires de chromosomes homologues. Cela est dû au fait que les chromosomes non homologues sont combinés dans la cellule indépendamment les uns des autres, ce qui a été prouvé non seulement par l'étude du caractère héréditaire des caractères, mais également par une méthode cytologique directe.
Lors de l'étude du matériel, faites attention aux cas de violations de la division régulière par phénotype causées par l'action létale de gènes individuels.
Hérédité et variabilité. L'hérédité et la variabilité sont les propriétés les plus importantes caractéristiques de tous les organismes vivants.
La variabilité héréditaire, ou génotypique, est subdivisée en combinatoire et mutationnelle.
La variabilité est appelée combinatoire, qui repose sur la formation de recombinaisons, c'est-à-dire de combinaisons de gènes que les parents n'avaient pas.
La variabilité combinatoire est basée sur la reproduction sexuée des organismes, à la suite de laquelle une grande variété de génotypes apparaît. Trois processus sont des sources pratiquement illimitées de variation génétique :
Divergence indépendante des chromosomes homologues dans la première division méiotique. C'est la combinaison indépendante des chromosomes dans la méiose qui est à la base de la troisième loi de G. Mendel. L'apparition de graines de pois verts lisses et jaunes ridés dans la deuxième génération du croisement de plantes avec des graines jaunes lisses et vertes ridées est un exemple de variabilité combinatoire.
Échange mutuel de régions de chromosomes homologues, ou croisement. Il crée de nouveaux groupes de liaison, c'est-à-dire qu'il sert de source importante de recombinaison génétique d'allèles. Une fois dans le zygote, les chromosomes recombinants contribuent à l'apparition de traits atypiques pour chacun des parents.
Combinaison aléatoire de gamètes pendant la fécondation.
Ces sources de variabilité combinatoire agissent indépendamment et simultanément, tout en assurant un « brassage » constant de gènes, ce qui conduit à l'émergence d'organismes de génotype et de phénotype différents (les gènes eux-mêmes ne changent pas). Cependant, les nouvelles combinaisons de gènes se décomposent assez facilement lorsqu'elles sont transmises de génération en génération.
Un exemple de variabilité combinatoire. La fleur de beauté nocturne a un gène pour les pétales rouges A et un gène pour le blanc a. Le corps d'Aa a des pétales roses. Ainsi, la beauté de la nuit n'a pas de gène rose ; le rose apparaît lorsqu'une combinaison (combinaison) d'un gène rouge et d'un blanc.
La personne a une maladie héréditaire drépanocytaire. AA est la norme, aa est la mort, Aa est SKA. Avec SKA, une personne ne peut pas tolérer un effort physique accru, alors qu'elle ne souffre pas de paludisme, c'est-à-dire que l'agent causal du paludisme Plasmodium malaria ne peut pas se nourrir de la mauvaise hémoglobine. Cette caractéristique est utile dans la ceinture équatoriale ; il n'y a pas de gène pour cela, il résulte d'une combinaison des gènes A et a.
Ainsi, la variation héréditaire est renforcée par la variation combinatoire. Une fois apparues, des mutations individuelles se trouvent à proximité d'autres mutations, font partie de nouveaux génotypes, c'est-à-dire que de nombreuses combinaisons d'allèles apparaissent. Tout individu est génétiquement unique (à l'exception des jumeaux identiques et des individus nés de la reproduction asexuée d'un clone avec une seule cellule comme ancêtre). Donc, si nous supposons que dans chaque paire de chromosomes homologues il n'y a qu'une seule paire de gènes alléliques, alors pour une personne dont l'ensemble haploïde de chromosomes est de 23, le nombre de génotypes possibles sera de 3 à 23 degrés. Un si grand nombre de génotypes est 20 fois le nombre de tous les habitants de la Terre. Cependant, en réalité, les chromosomes homologues diffèrent dans plusieurs gènes et le calcul ne prend pas en compte le phénomène de croisement. . Par conséquent, le nombre de génotypes possibles est exprimé en nombres astronomiques, et on peut affirmer avec certitude que la présence de deux personnes identiques est pratiquement improbable (à l'exception de jumeaux identiques, issus du même œuf fécondé). D'où, en particulier, la possibilité d'une identification fiable de la personne par les restes de tissus vivants, la confirmation ou l'exclusion de la paternité.
Ainsi, l'échange de gènes dû au croisement de chromosomes dans la première division de la méiose, la recombinaison indépendante et aléatoire des chromosomes dans la méiose, et la chance de fusion des gamètes dans le processus sexuel sont trois facteurs qui assurent l'existence d'une variabilité combinatoire. Variabilité mutationnelle du génotype lui-même.
Les mutations sont des modifications héréditaires soudaines du matériel génétique, entraînant une modification de certaines caractéristiques d'un organisme.
Les principales dispositions de la théorie mutationnelle ont été développées par le scientifique G. De Vries en 1901 – 1903 et résumer à ceci :
Les mutations apparaissent soudainement, à pas de géant, sous forme de changements discrets de traits ;
Contrairement aux changements non héréditaires, les mutations sont des changements qualitatifs qui se transmettent de génération en génération ;
Les mutations se manifestent de différentes manières et peuvent être à la fois bénéfiques et néfastes, dominantes et récessives ;
La probabilité de détecter des mutations dépend du nombre d'individus examinés ;
Des mutations similaires peuvent se reproduire ;
Les mutations sont non directionnelles (spontanées), c'est-à-dire que n'importe quelle partie du chromosome peut muter, provoquant des changements dans les signes mineurs et vitaux.
Presque tout changement dans la structure ou le nombre de chromosomes, dans lesquels la cellule conserve la capacité de se reproduire, provoque une modification héréditaire des caractéristiques de l'organisme.
Par la nature du changement dans le génome, c'est-à-dire l'ensemble de gènes contenus dans l'ensemble haploïde de chromosomes, on distingue les mutations génétiques, chromosomiques et génomiques.
Les mutations génétiques, ou ponctuelles, sont le résultat d'un changement dans la séquence nucléotidique d'une molécule d'ADN au sein d'un gène.
Une telle modification du gène est reproduite lors de la transcription dans la structure de l'i-ARN ; elle conduit à une modification de la séquence des acides aminés dans la chaîne polypeptidique formée lors de la traduction sur les ribosomes. En conséquence, une autre protéine est synthétisée, ce qui entraîne une modification de la caractéristique correspondante de l'organisme. C'est le type de mutation le plus courant et la source la plus importante de variation héréditaire dans les organismes.
Les mutations chromosomiques (réarrangements ou aberrations) sont des changements dans la structure des chromosomes qui peuvent être identifiés et étudiés au microscope optique.
Des réarrangements de différents types sont connus :
un manque de – perte des extrémités du chromosome;
Effacement – perte d'une partie du chromosome dans sa partie médiane;
Reproduction – deux - ou plusieurs répétitions de gènes localisés dans une certaine partie du chromosome;
Inversion – rotation de la région chromosomique de 180 °, à la suite de laquelle les gènes de cette région sont situés dans la séquence opposée à celle habituelle;
Déplacement – changer la position de n'importe quelle partie du chromosome dans l'ensemble de chromosomes. Le type de translocation le plus courant est la translocation réciproque, dans laquelle des zones sont échangées entre deux chromosomes non homologues. Une section d'un chromosome peut changer de position sans échange réciproque, rester dans le même chromosome ou être incluse dans un autre.
Mutations génomiques - une modification du nombre de chromosomes dans le génome des cellules d'un organisme. Ce phénomène se produit dans deux directions : vers une augmentation du nombre d'ensembles haploïdes entiers (polyploïdie) et vers la perte ou l'inclusion de chromosomes individuels (aneuploïdie).
Polyploïdie – une augmentation multiple de l'ensemble haploïde de chromosomes. Les cellules avec un nombre différent d'ensembles de chromosomes haploïdes sont appelées triploïdes (3 n), tétraploïdes (4 n), hexaploïdes (6 n), octaploïdes (8 n), etc. Le plus souvent, les polyploïdes se forment lorsque l'ordre de divergence chromosomique à les pôles cellulaires pendant la méiose sont perturbés ou mitoses. La polyploïdie entraîne une modification des caractéristiques d'un organisme et est donc une source importante de variabilité dans l'évolution et la sélection, en particulier chez les plantes. Cela est dû au fait que l'hermaphrodisme (autopollinisation), l'apomixie (parthénogenèse) et la reproduction végétative sont très répandus dans les organismes végétaux. Par conséquent, environ un tiers des espèces végétales communes sur notre planète, – polyploïdes, et dans les conditions fortement continentales du Pamir de haute montagne, jusqu'à 85% des polyploïdes se développent. Presque toutes les plantes cultivées sont également polyploïdes, qui, contrairement à leurs parents sauvages, ont des fleurs, des fruits et des graines plus gros, et plus de nutriments s'accumulent dans les organes de stockage (tige, tubercules). Les polyploïdes s'adaptent plus facilement aux conditions de vie défavorables, tolèrent plus facilement les basses températures et la sécheresse. C'est pourquoi ils sont répandus dans les régions du nord et de haute montagne.