Αποκρυπτογράφηση του γονιδιώματος του χιμπατζή και σύγκρισή του με το ανθρώπινο γονιδίωμα. Η Drosophila με ένα αρχαίο γονίδιο διέψευσε μια από τις κλασικές θεωρίες της εξέλιξης
Στην 50ή επέτειο από την ανακάλυψη της δομής του DNA
A.V. Ζελένιν
ΦΥΤΙΚΟ ΓΟΝΙΔΩΜΑ
A. V. Zelenin
Ζέλεν Αλεξάντερ Βλαντιμίροβιτς- Διδάκτωρ Βιολογικών Επιστημών,
Προϊστάμενος Εργαστηρίου, Ινστιτούτο Μοριακής Βιολογίας. V.A. Engelhardt RAS.
Τα εντυπωσιακά επιτεύγματα του προγράμματος του ανθρώπινου γονιδιώματος, καθώς και η επιτυχία των εργασιών για την αποκρυπτογράφηση των λεγόμενων εξαιρετικά μικρών (ιοί), μικρού μεγέθους (βακτήρια, ζυμομύκητες) και μεσαίου μεγέθους (στρογγυλός σκώληκας, Drosophila), κατέστησαν δυνατή την μεταβείτε σε μια μεγάλης κλίμακας μελέτη μεγάλων και πολύ μεγάλων φυτικών γονιδιωμάτων. Η επείγουσα ανάγκη για λεπτομερή μελέτη των γονιδιωμάτων των πιο σημαντικών από οικονομική άποψη φυτών τονίστηκε σε μια συνάντηση για τη γονιδιωματική των φυτών που πραγματοποιήθηκε το 1997 στις ΗΠΑ [,]. Με τα χρόνια από τότε, έχουν σημειωθεί αναμφισβήτητες επιτυχίες σε αυτόν τον τομέα. Το 2000, εμφανίστηκε μια δημοσίευση για την πλήρη αλληλούχιση (καθιέρωση της γραμμικής νουκλεοτιδικής αλληλουχίας όλου του πυρηνικού DNA) του γονιδιώματος της μικρής μουστάρδας - Arabidopsis, και το 2001 - για την προκαταρκτική (προσχέδιο) αλληλούχιση του γονιδιώματος του ρυζιού. Εργασίες για την αλληλούχιση μεγάλων και εξαιρετικά μεγάλων φυτικών γονιδιωμάτων (καλαμπόκι, σίκαλη, σιτάρι) έχουν αναφερθεί επανειλημμένα, αλλά αυτά τα μηνύματα δεν περιείχαν συγκεκριμένες πληροφορίες και ήταν μάλλον δηλώσεις προθέσεων.
Αναμένεται ότι η αποκρυπτογράφηση των γονιδιωμάτων των φυτών θα ανοίξει ευρείες προοπτικές για την επιστήμη και την πρακτική. Καταρχάς, ο εντοπισμός νέων γονιδίων και η αλυσίδα της γενετικής τους ρύθμισης θα αυξήσει σημαντικά την παραγωγικότητα των φυτών μέσω της χρήσης βιοτεχνολογικών προσεγγίσεων. Η ανακάλυψη, η απομόνωση, η αναπαραγωγή (κλωνοποίηση) και η αλληλουχία γονιδίων που είναι υπεύθυνα για σημαντικές λειτουργίες του φυτικού οργανισμού όπως η αναπαραγωγή και η παραγωγικότητα, οι διαδικασίες μεταβλητότητας, η αντοχή σε δυσμενείς περιβαλλοντικούς παράγοντες, καθώς και το ομόλογο ζευγάρωμα χρωμοσωμάτων, σχετίζεται με την εμφάνιση νέων ευκαιριών για τη βελτίωση της διαδικασίας επιλογής . Τέλος, απομονωμένα και κλωνοποιημένα γονίδια μπορούν να χρησιμοποιηθούν για τη λήψη διαγονιδιακών φυτών με θεμελιωδώς νέες ιδιότητες και την ανάλυση των μηχανισμών ρύθμισης της γονιδιακής δραστηριότητας.
Η σημασία της μελέτης του γονιδιώματος των φυτών τονίζεται επίσης από το γεγονός ότι μέχρι στιγμής ο αριθμός των εντοπισμένων, κλωνοποιημένων και αλληλουχούμενων γονιδίων φυτών είναι μικρός και, σύμφωνα με διάφορες εκτιμήσεις, κυμαίνεται μεταξύ 800 και 1200. Αυτός είναι 10-15 φορές μικρότερος από για παράδειγμα, στους ανθρώπους.
Οι Ηνωμένες Πολιτείες παραμένουν ο αναμφισβήτητος ηγέτης στη μεγάλης κλίμακας μελέτη των γονιδιωμάτων των φυτών, αν και εντατική έρευνα για το γονιδίωμα του ρυζιού διεξάγεται στην Ιαπωνία και τα τελευταία χρόνια στην Κίνα. Εκτός από τα εργαστήρια των ΗΠΑ, ευρωπαϊκές ερευνητικές ομάδες συμμετείχαν ενεργά στην αποκρυπτογράφηση του γονιδιώματος Arabidopsis. Η προφανής ηγεσία των Ηνωμένων Πολιτειών προκαλεί σοβαρή ανησυχία μεταξύ των Ευρωπαίων επιστημόνων, την οποία εξέφρασαν ξεκάθαρα σε μια συνάντηση με νόημα με τίτλο «Προοπτικές για τη Γονιδιωματική στη Μεταγονιδιωματική Εποχή», που πραγματοποιήθηκε στη Γαλλία στα τέλη του 2000. Η πρόοδος της αμερικανικής επιστήμης στη μελέτη των γονιδιωμάτων των γεωργικών φυτών και στη δημιουργία διαγονιδιακών μορφών φυτών, σύμφωνα με Ευρωπαίους επιστήμονες, απειλεί ότι στο όχι πολύ μακρινό μέλλον (από δύο έως πέντε δεκαετίες), όταν η αύξηση του πληθυσμού θα φέρει την ανθρωπότητα αντιμέτωπη με η γενική επισιτιστική κρίση, η ευρωπαϊκή οικονομία και η επιστήμη θα εξαρτηθούν από την αμερικανική τεχνολογία. Στο πλαίσιο αυτό, ανακοινώθηκε η δημιουργία γαλλογερμανικού επιστημονικού προγράμματος για τη μελέτη των γονιδιωμάτων των φυτών (Plantgene) και η επένδυση σημαντικών κεφαλαίων σε αυτό.
Προφανώς, τα προβλήματα της γονιδιωματικής των φυτών πρέπει να προσελκύσουν την προσοχή των Ρώσων επιστημόνων και οργανωτών επιστήμης, καθώς και των κυβερνώντων οργάνων, καθώς μιλάμε όχι μόνο για επιστημονικό κύρος, αλλά και για την εθνική ασφάλεια της χώρας. Σε μία ή δύο δεκαετίες, τα τρόφιμα θα γίνουν ο πιο σημαντικός στρατηγικός πόρος.
ΔΥΣΚΟΛΙΕΣ ΣΤΗ ΜΕΛΕΤΗ ΤΩΝ ΦΥΤΩΝ ΓΟΝΙΔΙΩΜΑΤΩΝ
Η μελέτη του γονιδιώματος των φυτών είναι μια πολύ πιο σύνθετη εργασία από τη μελέτη του γονιδιώματος των ανθρώπων και άλλων ζώων. Αυτό οφείλεται στις ακόλουθες συνθήκες:
τεράστια μεγέθη γονιδιώματος, που φτάνουν δεκάδες, ακόμη και εκατοντάδες δισεκατομμύρια ζεύγη νουκλεοτιδίων (bp) για μεμονωμένα φυτικά είδη: τα γονιδιώματα των κύριων οικονομικά σημαντικών φυτών (εκτός από το ρύζι, το λινάρι και το βαμβάκι) είναι είτε κοντά σε μέγεθος με το ανθρώπινο γονιδίωμα είτε το υπερβαίνουν πολλές φορές (πίνακας)?ΜΕΛΕΤΕΣ ΧΡΩΜΟΣΩΜΙΚΟΥ ΓΟΝΙΔΩΜΑΤΟΣΑπότομες διακυμάνσεις στον αριθμό των χρωμοσωμάτων σε διαφορετικά φυτά - από δύο σε ορισμένα είδη έως αρκετές εκατοντάδες σε άλλα, και δεν είναι δυνατό να εντοπιστεί μια αυστηρή συσχέτιση μεταξύ του μεγέθους του γονιδιώματος και του αριθμού των χρωμοσωμάτων.
Μια αφθονία πολυπλοειδών (που περιέχει περισσότερα από δύο γονιδιώματα ανά κύτταρο) σχηματίζεται με παρόμοια αλλά όχι πανομοιότυπα γονιδιώματα (αλλοπολυπλοειδία).
Ο ακραίος εμπλουτισμός των φυτικών γονιδιωμάτων (έως 99%) με «ασήμαντο» (μη κωδικοποιητικό, δηλαδή που δεν περιέχει γονίδια) DNA, το οποίο περιπλέκει πολύ την ένωση (διάταξη με τη σωστή σειρά) των αλληλουχούμενων θραυσμάτων σε ένα κοινό μεγάλο περιοχή DNA μεγέθους (contig);
Ελλιπής (σε σύγκριση με τα γονιδιώματα της Drosophila, του ανθρώπου και του ποντικού) μορφολογική, γενετική και φυσική χαρτογράφηση των χρωμοσωμάτων.
Η πρακτική αδυναμία απομόνωσης μεμονωμένων χρωμοσωμάτων σε καθαρή μορφή με τη χρήση μεθόδων που χρησιμοποιούνται συνήθως για το σκοπό αυτό για ανθρώπινα και ζωικά χρωμοσώματα (διαλογή ροής και χρήση υβριδίων κυττάρων).
Η δυσκολία της χρωμοσωμικής χαρτογράφησης (καθορισμός της θέσης στο χρωμόσωμα) μεμονωμένων γονιδίων με χρήση υβριδισμού επί τόπου, λόγω τόσο της υψηλής περιεκτικότητας σε «ασήμαντο» DNA στα γονιδιώματα των φυτών και των ιδιαιτεροτήτων της δομικής οργάνωσης των φυτικών χρωμοσωμάτων.
Η εξελικτική απόσταση των φυτών από τα ζώα, η οποία περιπλέκει σοβαρά τη χρήση των πληροφοριών που λαμβάνονται από την αλληλουχία του γονιδιώματος των ανθρώπων και άλλων ζώων για τη μελέτη γονιδιωμάτων φυτών.
Η μακρά διαδικασία αναπαραγωγής των περισσότερων φυτών, που επιβραδύνει σημαντικά τη γενετική τους ανάλυση.
Οι χρωμοσωμικές (κυτταρογενετικές) μελέτες των γονιδιωμάτων γενικά και των φυτών ειδικότερα έχουν μακρά ιστορία. Ο όρος «γονιδίωμα» προτάθηκε για να δηλώσει ένα απλοειδές (μονό) σύνολο χρωμοσωμάτων με τα γονίδια που περιέχουν στο πρώτο τέταρτο του 20ου αιώνα, δηλαδή πολύ πριν καθιερωθεί ο ρόλος του DNA ως φορέα γενετικής πληροφορίας.
Η περιγραφή του γονιδιώματος ενός νέου, προηγουμένως μη μελετημένου γενετικά πολυκύτταρου οργανισμού ξεκινά συνήθως με τη μελέτη και περιγραφή του πλήρους συνόλου των χρωμοσωμάτων του (καρυότυπος). Αυτό φυσικά ισχύει και για τα φυτά, ένας τεράστιος αριθμός των οποίων δεν έχει καν αρχίσει να μελετάται.
Ήδη στην αυγή των χρωμοσωμικών μελετών, γονιδιώματα συγγενών φυτικών ειδών συγκρίθηκαν με βάση την ανάλυση της μειωτικής σύζευξης (ενοποίηση ομόλογων χρωμοσωμάτων) σε μεσοειδικά υβρίδια. Τα τελευταία 100 χρόνια, οι δυνατότητες της χρωμοσωμικής ανάλυσης έχουν επεκταθεί δραματικά. Σήμερα, χρησιμοποιούνται πιο προηγμένες τεχνολογίες για τον χαρακτηρισμό των γονιδιωμάτων των φυτών: διάφορες παραλλαγές της λεγόμενης διαφορικής χρώσης, που καθιστά δυνατό τον εντοπισμό μεμονωμένων χρωμοσωμάτων με βάση μορφολογικά χαρακτηριστικά. παραγωγή μικτών γενών επί τόπου,καθιστώντας δυνατό τον εντοπισμό συγκεκριμένων γονιδίων στα χρωμοσώματα· βιοχημικές μελέτες κυτταρικών πρωτεϊνών (ηλεκτροφόρηση και ανοσοχημεία) και, τέλος, ένα σύνολο μεθόδων που βασίζονται στην ανάλυση του χρωμοσωμικού DNA μέχρι τον προσδιορισμό της αλληλουχίας του.

Ρύζι. 1.Καρυότυποι δημητριακών: α - σίκαλη (14 χρωμοσώματα), β - σκληρό σιτάρι (28 χρωμοσώματα), γ - μαλακό σιτάρι (42 χρωμοσώματα), δ - κριθάρι (14 χρωμοσώματα)Οι καρυότυποι των δημητριακών, κυρίως του σιταριού και της σίκαλης, έχουν μελετηθεί εδώ και πολλά χρόνια. Είναι ενδιαφέρον ότι σε διαφορετικά είδη αυτών των φυτών ο αριθμός των χρωμοσωμάτων είναι διαφορετικός, αλλά πάντα πολλαπλάσιο του επτά. Τα μεμονωμένα είδη δημητριακών μπορούν να αναγνωριστούν αξιόπιστα από τον καρυότυπο τους. Για παράδειγμα, το γονιδίωμα της σίκαλης αποτελείται από επτά ζεύγη μεγάλων χρωμοσωμάτων με έντονα χρωματισμένα ετεροχρωματικά μπλοκ στα άκρα τους, που συχνά ονομάζονται τμήματα ή ζώνες (Εικ. 1α). Τα γονιδιώματα του σίτου έχουν ήδη 14 και 21 ζεύγη χρωμοσωμάτων (Εικ. 1, β, γ) και η κατανομή των ετεροχρωματικών μπλοκ σε αυτά δεν είναι η ίδια όπως στα χρωμοσώματα της σίκαλης. Τα επιμέρους γονιδιώματα του σιταριού, που ονομάζονται A, B και D, διαφέρουν επίσης μεταξύ τους. Η αύξηση του αριθμού των χρωμοσωμάτων από 14 σε 21 οδηγεί σε μια απότομη αλλαγή στις ιδιότητες του σίτου, η οποία αντικατοπτρίζεται στα ονόματά τους: σκληρό, ή μακαρόνια, σιτάρι και μαλακό, ή ψωμί, σιτάρι . Το γονίδιο D, το οποίο περιέχει γονίδια για πρωτεΐνες γλουτένης, είναι υπεύθυνο για την απόκτηση υψηλών ιδιοτήτων ψησίματος από το μαλακό σιτάρι, το οποίο δίνει στη ζύμη τη λεγόμενη βλάστηση. Σε αυτό το γονιδίωμα δίνεται ιδιαίτερη προσοχή στη βελτίωση της επιλογής του ψωμιού. Ένα άλλο δημητριακό 14 χρωμοσωμάτων, το κριθάρι (Εικ. 1, δ), δεν χρησιμοποιείται συνήθως για την παρασκευή ψωμιού, αλλά χρησιμεύει ως η κύρια πρώτη ύλη για την παραγωγή κοινών προϊόντων όπως η μπύρα και το ουίσκι.
Μελετώνται εντατικά τα χρωμοσώματα ορισμένων άγριων φυτών που χρησιμοποιούνται για τη βελτίωση της ποιότητας των σημαντικότερων γεωργικών ειδών, για παράδειγμα των άγριων συγγενών του σιταριού - Αιγίλωπα. Δημιουργούνται νέες μορφές φυτών μέσω διασταύρωσης (Εικ. 2) και επιλογής. Τα τελευταία χρόνια, σημαντικές βελτιώσεις στις ερευνητικές μεθόδους κατέστησαν δυνατή την έναρξη της μελέτης των γονιδιωμάτων των φυτών των οποίων τα χαρακτηριστικά καρυότυπου (κυρίως μικρά μεγέθη χρωμοσωμάτων) τα καθιστούσαν προηγουμένως απρόσιτα για χρωμοσωμική ανάλυση. Έτσι, μόλις πρόσφατα αναγνωρίστηκαν για πρώτη φορά όλα τα χρωμοσώματα του βαμβακιού, του χαμομηλιού και του λιναριού.

Ρύζι. 2. Καρυότυποι σιταριού και υβρίδιο σιταριού-Αιγίλωψ
α - εξαπλοειδές μαλακό σιτάρι ( Triticum astivum), που αποτελείται από γονιδιώματα Α, Β και Ο. β - τετραπλοειδές σιτάρι ( Triticum timopheevi), που αποτελείται από γονιδιώματα Α και G. περιέχει γονίδια για αντοχή στις περισσότερες ασθένειες του σιταριού. γ - υβρίδια Triticum astivumΧ Triticum timopheevi, ανθεκτικό στο ωίδιο και τη σκουριά, η αντικατάσταση μέρους των χρωμοσωμάτων είναι σαφώς ορατήΠΡΩΤΟΓΕΝΙΚΗ ΔΟΜΗ ΤΟΥ DNA
Καθώς η μοριακή γενετική αναπτύχθηκε, η ίδια η έννοια του γονιδιώματος διευρύνθηκε. Τώρα αυτός ο όρος ερμηνεύεται τόσο με την κλασική χρωμοσωμική όσο και με τη σύγχρονη μοριακή έννοια: ολόκληρο το γενετικό υλικό ενός μεμονωμένου ιού, κυττάρου και οργανισμού. Φυσικά, μετά τη μελέτη της πλήρους πρωτογενούς δομής των γονιδιωμάτων (όπως αποκαλείται συχνά η πλήρης γραμμική ακολουθία βάσεων νουκλεϊκών οξέων) ορισμένων μικροοργανισμών και ανθρώπων, προέκυψε το ζήτημα της αλληλουχίας των γονιδιωμάτων των φυτών.
Από τους πολλούς φυτικούς οργανισμούς, δύο επιλέχθηκαν για μελέτη - το Arabidopsis, που αντιπροσωπεύει την κατηγορία των δικοτυληδόνων (μέγεθος γονιδιώματος 125 εκατομμύρια bp) και το ρύζι από την κατηγορία των μονοκοτυληδόνων (420-470 εκατομμύρια bp). Αυτά τα γονιδιώματα είναι μικρά σε σύγκριση με άλλα φυτικά γονιδιώματα και περιέχουν σχετικά λίγες επαναλαμβανόμενες τομές DNA. Τέτοια χαρακτηριστικά έδιναν ελπίδα ότι τα επιλεγμένα γονιδιώματα θα ήταν προσβάσιμα για σχετικά γρήγορο προσδιορισμό της πρωτογενούς δομής τους.

Ρύζι. 3. Arabidopsis - μικρή μουστάρδα - ένα μικρό φυτό από την οικογένεια των σταυρανθών ( Brassicaceae). Σε χώρο ίσο σε έκταση με μία σελίδα του περιοδικού μας, μπορούν να αναπτυχθούν έως και χίλιοι μεμονωμένοι οργανισμοί ArabidopsisΗ βάση για την επιλογή του Arabidopsis δεν ήταν μόνο το μικρό μέγεθος του γονιδιώματός του, αλλά και το μικρό μέγεθος του οργανισμού, γεγονός που καθιστά εύκολη την ανάπτυξή του σε εργαστηριακές συνθήκες (Εικ. 3). Λάβαμε υπόψη τον σύντομο αναπαραγωγικό του κύκλο, χάρη στον οποίο είναι δυνατή η γρήγορη διεξαγωγή πειραμάτων διασταύρωσης και επιλογής, λεπτομερής γενετική, ευκολία χειρισμού με μεταβαλλόμενες συνθήκες καλλιέργειας (αλλαγή της σύνθεσης αλατιού του εδάφους, προσθήκη διαφορετικών θρεπτικών συστατικών κ.λπ.) και έλεγχος της επίδρασης στα φυτά διαφόρων μεταλλαξογόνων παραγόντων και παθογόνων (ιοί, βακτήρια, μύκητες). Το Arabidopsis δεν έχει οικονομική αξία, επομένως το γονιδίωμά του, μαζί με το γονιδίωμα του ποντικιού, ονομάστηκε γονιδίωμα αναφοράς, ή, με μικρότερη ακρίβεια, μοντέλο γονιδιώματος.*
* Η εμφάνιση του όρου «μοντέλο γονιδιώματος» στη ρωσική βιβλιογραφία είναι το αποτέλεσμα μιας ανακριβούς μετάφρασης της αγγλικής φράσης μοντέλο γονιδιώματος. Η λέξη «μοντέλο» σημαίνει όχι μόνο το επίθετο «μοντέλο», αλλά και το ουσιαστικό «δείγμα», «πρότυπο», «μοντέλο». Θα ήταν πιο σωστό να μιλήσουμε για ένα δείγμα γονιδιώματος ή ένα γονιδίωμα αναφοράς.Η εντατική εργασία για τον προσδιορισμό της αλληλουχίας του γονιδιώματος Arabidopsis ξεκίνησε το 1996 από μια διεθνή κοινοπραξία που περιελάμβανε επιστημονικά ιδρύματα και ερευνητικές ομάδες από τις ΗΠΑ, την Ιαπωνία, το Βέλγιο, την Ιταλία, τη Μεγάλη Βρετανία και τη Γερμανία. Τον Δεκέμβριο του 2000, έγιναν διαθέσιμες εκτενείς πληροφορίες που συνοψίζουν τον προσδιορισμό της πρωτογενούς δομής του γονιδιώματος Arabidopsis. Για την αλληλουχία χρησιμοποιήσαμε κλασική ή ιεραρχική τεχνολογία: πρώτα μελετήθηκαν μεμονωμένες μικρές τομές του γονιδιώματος, από τις οποίες κατασκευάστηκαν μεγαλύτερες τομές (contigs) και στο τελικό στάδιο, η δομή των μεμονωμένων χρωμοσωμάτων. Το πυρηνικό DNA του γονιδιώματος Arabidopsis κατανέμεται σε πέντε χρωμοσώματα. Το 1999 δημοσιεύθηκαν τα αποτελέσματα της αλληλούχισης δύο χρωμοσωμάτων και η δημοσίευση πληροφοριών σχετικά με την πρωτογενή δομή των υπόλοιπων τριών ολοκλήρωσε την αλληλούχιση ολόκληρου του γονιδιώματος.
Από 125 εκατομμύρια ζεύγη νουκλεοτιδίων, έχει προσδιοριστεί η πρωτογενής δομή των 119 εκατομμυρίων, που είναι το 92% ολόκληρου του γονιδιώματος. Μόνο το 8% του γονιδιώματος Arabidopsis, που περιέχει μεγάλα μπλοκ επαναλαμβανόμενων τμημάτων DNA, αποδείχθηκε ότι δεν ήταν προσβάσιμο για μελέτη. Όσον αφορά την πληρότητα και την πληρότητα της αλληλουχίας των ευκαρυωτικών γονιδιωμάτων, το Arabidopsis παραμένει στους τρεις πρώτους πρωταθλητές μαζί με τον μονοκύτταρο οργανισμό ζύμης Saccharomyces cerevisiaeκαι πολυκύτταρος ζωικός οργανισμός Caenorhabditis κομψότητα(βλέπε πίνακα).
Περίπου 15 χιλιάδες μεμονωμένα γονίδια που κωδικοποιούν πρωτεΐνες βρέθηκαν στο γονιδίωμα Arabidopsis. Περίπου 12 χιλιάδες από αυτά περιέχονται σε δύο αντίγραφα ανά απλοειδές (μονό) γονιδίωμα, επομένως ο συνολικός αριθμός γονιδίων είναι 27 χιλιάδες. Ο αριθμός των γονιδίων στο Arabidopsis δεν διαφέρει πολύ από τον αριθμό των γονιδίων σε οργανισμούς όπως ο άνθρωπος και τα ποντίκια. αλλά το μέγεθος του γονιδιώματός του 25-30 φορές μικρότερο. Αυτή η περίσταση συνδέεται με σημαντικά χαρακτηριστικά στη δομή των μεμονωμένων γονιδίων Arabidopsis και στη συνολική δομή του γονιδιώματός του.
Τα γονίδια Arabidopsis είναι συμπαγή, περιέχουν μόνο λίγα εξόνια (περιοχές κωδικοποίησης πρωτεΐνης), που διαχωρίζονται από μικρές (περίπου 250 bp) μη κωδικοποιητικές τεντώσεις DNA (εσώνια). Τα κενά μεταξύ των μεμονωμένων γονιδίων είναι κατά μέσο όρο 4,6 χιλιάδες ζεύγη νουκλεοτιδίων. Για σύγκριση, επισημαίνουμε ότι τα ανθρώπινα γονίδια περιέχουν πολλές δεκάδες, ακόμη και εκατοντάδες εξόνια και ιντρόνια, και οι διαγονιδιακές περιοχές έχουν μεγέθη 10 χιλιάδων ζευγών νουκλεοτιδίων ή περισσότερο. Πιστεύεται ότι η παρουσία ενός μικρού συμπαγούς γονιδιώματος συνέβαλε στην εξελικτική σταθερότητα του Arabidopsis, καθώς το DNA του έγινε λιγότερο στόχος για διάφορους επιβλαβείς παράγοντες, ιδίως για την εισαγωγή επαναλαμβανόμενων θραυσμάτων DNA (τρανσποζόνια) που μοιάζουν με ιούς στο γονιδίωμα.
Άλλα μοριακά χαρακτηριστικά του γονιδιώματος Arabidopsis περιλαμβάνουν τον εμπλουτισμό εξονίων με γουανίνη και κυτοσίνη (44% σε εξόνια και 32% σε ιντρόνια) σε σύγκριση με τα ζωικά γονίδια, καθώς και την παρουσία διπλών επαναλαμβανόμενων (διπλασιασμένων) γονιδίων. Πιστεύεται ότι αυτός ο διπλασιασμός προέκυψε ως αποτέλεσμα τεσσάρων ταυτόχρονων γεγονότων, τα οποία συνίσταντο στον διπλασιασμό (επανάληψη) μέρους των γονιδίων Arabidopsis ή τη σύντηξη σχετικών γονιδιωμάτων. Αυτά τα γεγονότα, που έλαβαν χώρα πριν από 100-200 εκατομμύρια χρόνια, είναι μια εκδήλωση της γενικής τάσης προς πολυπλοειδισμό (πολλαπλή αύξηση του αριθμού των γονιδιωμάτων σε έναν οργανισμό), χαρακτηριστικό των φυτικών γονιδιωμάτων. Ωστόσο, ορισμένα γεγονότα δείχνουν ότι στο Arabidopsis τα διπλά γονίδια δεν είναι πανομοιότυπα και λειτουργούν διαφορετικά, κάτι που μπορεί να οφείλεται σε μεταλλάξεις στις ρυθμιστικές τους περιοχές.
Ένα άλλο αντικείμενο πλήρους προσδιορισμού αλληλουχίας DNA ήταν το ρύζι. Το γονιδίωμα αυτού του φυτού είναι επίσης μικρό (12 χρωμοσώματα, που δίνουν συνολικά 420-470 εκατομμύρια bp), μόλις 3,5 φορές μεγαλύτερο από αυτό του Arabidopsis. Ωστόσο, σε αντίθεση με το Arabidopsis, το ρύζι έχει τεράστια οικονομική σημασία, καθώς αποτελεί τη βάση της διατροφής για περισσότερο από το ήμισυ της ανθρωπότητας, επομένως όχι μόνο δισεκατομμύρια καταναλωτές ενδιαφέρονται ζωτικά για τη βελτίωση των ιδιοτήτων του, αλλά και ένας στρατός πολλών εκατομμυρίων δολαρίων ανθρώπων που συμμετέχουν ενεργά η πολύ κοπιαστική διαδικασία ανάπτυξής του.
Μερικοί ερευνητές άρχισαν να μελετούν το γονιδίωμα του ρυζιού στη δεκαετία του '80 του περασμένου αιώνα, αλλά αυτή η εργασία έφτασε σε σοβαρή κλίμακα μόνο στη δεκαετία του '90. Το 1991, ένα πρόγραμμα για την αποκρυπτογράφηση της δομής του γονιδιώματος του ρυζιού δημιουργήθηκε στην Ιαπωνία, συνδυάζοντας τις προσπάθειες πολλών ερευνητικών ομάδων. Το 1997, με βάση αυτό το πρόγραμμα, οργανώθηκε το International Rice Genome Project. Οι συμμετέχοντες αποφάσισαν να επικεντρώσουν τις προσπάθειές τους στον προσδιορισμό της αλληλουχίας ενός από τα υποείδη ρυζιού ( Oriza sativajaponica), στη μελέτη της οποίας είχε ήδη σημειωθεί σημαντική πρόοδος μέχρι τότε. Το πρόγραμμα του ανθρώπινου γονιδιώματος έγινε σοβαρό κίνητρο και, μεταφορικά μιλώντας, αστέρι καθοδήγησης για μια τέτοια δουλειά.
Στο πλαίσιο αυτού του προγράμματος, δοκιμάστηκε η στρατηγική της «χρωμοσωμικής» ιεραρχικής διαίρεσης του γονιδιώματος, την οποία χρησιμοποίησαν οι συμμετέχοντες στη διεθνή κοινοπραξία για να αποκρυπτογραφήσουν το γονιδίωμα του ρυζιού. Ωστόσο, εάν, κατά τη μελέτη του ανθρώπινου γονιδιώματος, κλάσματα μεμονωμένων χρωμοσωμάτων απομονώθηκαν χρησιμοποιώντας διάφορες τεχνικές, τότε υλικό ειδικό για μεμονωμένα χρωμοσώματα ρυζιού και τα μεμονωμένα τμήματα τους ελήφθη με μικροτομή λέιζερ (αποκοπή μικροσκοπικών αντικειμένων). Στη διαφάνεια του μικροσκοπίου όπου βρίσκονται τα χρωμοσώματα του ρυζιού, υπό την επίδραση μιας δέσμης λέιζερ, όλα εκτός από το χρωμόσωμα ή τα τμήματα του που προορίζονται για ανάλυση καίγονται. Το υπόλοιπο υλικό χρησιμοποιείται για κλωνοποίηση και αλληλούχιση.
Έχουν δημοσιευθεί πολυάριθμες αναφορές σχετικά με τα αποτελέσματα της αλληλούχισης μεμονωμένων θραυσμάτων του γονιδιώματος του ρυζιού, που πραγματοποιήθηκαν με υψηλή ακρίβεια και λεπτομέρεια χαρακτηριστική της ιεραρχικής τεχνολογίας. Θεωρήθηκε ότι ο προσδιορισμός της πλήρους πρωτογενούς δομής του γονιδιώματος του ρυζιού θα ολοκληρωνόταν στα τέλη του 2003-μέσα του 2004 και τα αποτελέσματα, μαζί με δεδομένα για την πρωτογενή δομή του γονιδιώματος Arabidopsis, θα χρησιμοποιούνταν ευρέως στη συγκριτική γονιδιωματική άλλων φυτών.
Ωστόσο, στις αρχές του 2002, δύο ερευνητικές ομάδες - η μία από την Κίνα, η άλλη από την Ελβετία και τις Ηνωμένες Πολιτείες - δημοσίευσαν τα αποτελέσματα της πλήρους πρόχειρης (ακατέργαστης) αλληλουχίας του γονιδιώματος του ρυζιού, που πραγματοποιήθηκε με χρήση τεχνολογίας ολικής κλωνοποίησης. Σε αντίθεση με μια βαθμιαία (ιεραρχική) μελέτη, η συνολική προσέγγιση βασίζεται στην ταυτόχρονη κλωνοποίηση ολόκληρου του γονιδιωματικού DNA σε έναν από τους ιικούς ή βακτηριακούς φορείς και στη λήψη ενός σημαντικού (τεράστιου για μεσαία και μεγάλα γονιδιώματα) αριθμού μεμονωμένους κλώνους που περιέχουν διαφορετικά τμήματα DNA. Με βάση την ανάλυση αυτών των τμημάτων αλληλουχίας και την επικάλυψη πανομοιότυπων τελικών τμημάτων DNA, σχηματίζεται ένα contig - μια αλυσίδα αλληλουχιών DNA που ενώνονται μεταξύ τους. Το γενικό (ολικό) contig αντιπροσωπεύει την πρωτογενή δομή ολόκληρου του γονιδιώματος ή, τουλάχιστον, ενός μεμονωμένου χρωμοσώματος.
Σε μια τέτοια σχηματική παρουσίαση, η στρατηγική της συνολικής κλωνοποίησης φαίνεται απλή. Στην πραγματικότητα, αντιμετωπίζει σοβαρές δυσκολίες που σχετίζονται με την ανάγκη απόκτησης τεράστιου αριθμού κλώνων (είναι γενικά αποδεκτό ότι το γονιδίωμα ή η περιοχή του που μελετάται πρέπει να επικαλύπτεται από κλώνους τουλάχιστον 10 φορές), έναν τεράστιο όγκο αλληλουχίας και την εξαιρετικά πολύπλοκη εργασία σύνδεσης κλώνων, η οποία απαιτεί τη συμμετοχή ειδικών βιοπληροφορικής. Ένα σοβαρό εμπόδιο στην ολική κλωνοποίηση είναι η ποικιλία των επαναλαμβανόμενων περιοχών DNA, ο αριθμός των οποίων, όπως ήδη αναφέρθηκε, αυξάνεται απότομα καθώς αυξάνεται το μέγεθος του γονιδιώματος. Επομένως, η στρατηγική συνολικής αλληλουχίας χρησιμοποιείται κυρίως για τη μελέτη των γονιδιωμάτων ιών και μικροοργανισμών, αν και εφαρμόστηκε με επιτυχία για τη μελέτη του γονιδιώματος ενός πολυκύτταρου οργανισμού, του Drosophila.
Τα αποτελέσματα της συνολικής αλληλουχίας αυτού του γονιδιώματος «επιτέθηκαν» σε μια τεράστια σειρά πληροφοριών σχετικά με τη χρωμοσωμική, γονιδιακή και μοριακή του δομή που ελήφθησαν σε μια περίοδο σχεδόν 100 ετών μελέτης της Drosophila. Και όμως, όσον αφορά τον βαθμό αλληλουχίας, το γονιδίωμα Drosophila (66% του συνολικού μεγέθους του γονιδιώματος) είναι σημαντικά κατώτερο από το γονιδίωμα Arabidopsis (92%), παρά τα αρκετά παρόμοια μεγέθη τους - 180 εκατομμύρια και 125 εκατομμύρια ζεύγη νουκλεοτιδίων, αντίστοιχα. . Ως εκ τούτου, πρόσφατα προτάθηκε να ονομαστεί η τεχνολογία που χρησιμοποιείται για την αλληλουχία του γονιδιώματος της Drosophila μικτή.
Για την αλληλουχία του γονιδιώματος του ρυζιού, οι προαναφερθείσες ερευνητικές ομάδες πήραν δύο από τα υποείδη του, το πιο ευρέως καλλιεργούμενο στις ασιατικές χώρες - Oriza saliva L. ssp indicajΚαι Oriza saliva L. sspjaponica.Τα αποτελέσματα της έρευνάς τους συμπίπτουν από πολλές απόψεις, αλλά και διαφέρουν από πολλές απόψεις. Έτσι, εκπρόσωποι και των δύο ομάδων δήλωσαν ότι πέτυχαν αλληλεπικάλυψη περίπου 92-93% του γονιδιώματος. Έχει αποδειχθεί ότι περίπου το 42% του γονιδιώματος του ρυζιού αντιπροσωπεύεται από σύντομες επαναλήψεις DNA που αποτελούνται από 20 ζεύγη νουκλεοτιδίων και η πλειοψηφία των κινητών στοιχείων DNA (τρανσποζόνια) βρίσκονται σε διαγονιδιακές περιοχές. Ωστόσο, οι πληροφορίες σχετικά με το μέγεθος του γονιδιώματος του ρυζιού ποικίλλουν σημαντικά.
Για το ιαπωνικό υποείδος, το μέγεθος του γονιδιώματος προσδιορίζεται σε 466 εκατομμύρια ζεύγη νουκλεοτιδίων και για το ινδικό υποείδος - 420 εκατομμύρια. Ο λόγος αυτής της διαφοράς δεν είναι σαφής. Μπορεί να είναι συνέπεια διαφορετικών μεθοδολογικών προσεγγίσεων για τον προσδιορισμό του μεγέθους του μη κωδικοποιητικού μέρους των γονιδιωμάτων, δηλαδή, μπορεί να μην αντικατοπτρίζει την πραγματική κατάσταση πραγμάτων. Αλλά είναι πιθανό να υπάρχει πραγματικά μια διαφορά 15% στο μέγεθος των μελετηθέντων γονιδιωμάτων.
Η δεύτερη σοβαρή απόκλιση αποκαλύφθηκε στον αριθμό των γονιδίων που ανιχνεύθηκαν: για το ιαπωνικό υποείδος - από 46.022 έως 55.615 γονίδια ανά γονιδίωμα και για το ινδικό υποείδος - από 32.000 έως 50.000. Ο λόγος αυτής της διαφοράς δεν είναι σαφής.
Η ελλιπής και η ασυνέπεια των πληροφοριών που λαμβάνονται σημειώνεται στα σχόλια των δημοσιευμένων άρθρων. Ελπίζεται επίσης ότι τα κενά στη γνώση του γονιδιώματος του ρυζιού θα εξαλειφθούν συγκρίνοντας τα δεδομένα από την «πρόχειρη αλληλουχία» με τα αποτελέσματα της λεπτομερούς, ιεραρχικής αλληλουχίας που πραγματοποιήθηκε από συμμετέχοντες στο Διεθνές Πρόγραμμα Γονιδιώματος Ρυζιού.
ΣΥΓΚΡΙΤΙΚΗ ΚΑΙ ΛΕΙΤΟΥΡΓΙΚΗ ΓΟΝΙΔΙΩΜΑΤΙΚΗ ΦΥΤΩΝ
Τα εκτεταμένα δεδομένα που ελήφθησαν, τα μισά από τα οποία (τα αποτελέσματα της κινεζικής ομάδας) είναι διαθέσιμα στο κοινό, αναμφίβολα ανοίγουν ευρείες προοπτικές τόσο για τη μελέτη του γονιδιώματος του ρυζιού όσο και για τη γονιδιωματική των φυτών γενικότερα. Μια σύγκριση των ιδιοτήτων του γονιδιώματος Arabidopsis και ρυζιού έδειξε ότι τα περισσότερα από τα γονίδια (έως 80%) που προσδιορίζονται στο γονιδίωμα του Arabidopsis βρίσκονται επίσης στο γονιδίωμα του ρυζιού, ωστόσο, για περίπου τα μισά από τα γονίδια που βρίσκονται στο ρύζι, τα ανάλογα ( ορθολόγοι) δεν έχουν ακόμη βρεθεί στο γονιδίωμα Arabidopsis. Ταυτόχρονα, το 98% των γονιδίων των οποίων η πρωτογενής δομή έχει εδραιωθεί για άλλα δημητριακά έχει εντοπιστεί στο γονιδίωμα του ρυζιού.
Η σημαντική (σχεδόν διπλή) απόκλιση στον αριθμό των γονιδίων στο ρύζι και στο Arabidopsis προκαλεί σύγχυση. Ταυτόχρονα, τα δεδομένα από το πρόχειρο αντίγραφο του γονιδιώματος του ρυζιού, που ελήφθησαν με τη χρήση ολικής αλληλουχίας, πρακτικά δεν συγκρίνονται με τα εκτεταμένα αποτελέσματα της μελέτης του γονιδιώματος του ρυζιού χρησιμοποιώντας τη μέθοδο της ιεραρχικής κλωνοποίησης και αλληλούχισης, δηλαδή αυτό που έχει γίνει για το γονιδίωμα της Drosophila δεν έχει επιτευχθεί. Επομένως, παραμένει ασαφές εάν η διαφορά στον αριθμό των γονιδίων στο Arabidopsis και το ρύζι αντανακλά την πραγματική κατάσταση πραγμάτων ή εξηγείται από διαφορές στις μεθοδολογικές προσεγγίσεις.
Σε αντίθεση με το γονιδίωμα Arabidopsis, δεν παρέχονται πληροφορίες σχετικά με τα δίδυμα γονίδια στο γονιδίωμα του ρυζιού. Είναι πιθανό η σχετική αφθονία τους να είναι μεγαλύτερη στο ρύζι από ότι στο Arabidopsis. Αυτή η πιθανότητα υποστηρίζεται έμμεσα από στοιχεία για την παρουσία πολυπλοειδών μορφών ρυζιού. Μεγαλύτερη σαφήνεια σε αυτό το ζήτημα μπορεί να αναμένεται μετά την ολοκλήρωση του International Rice Genome Project και τη λήψη μιας λεπτομερούς εικόνας της πρωτογενούς δομής DNA αυτού του γονιδιώματος. Σοβαροί λόγοι για μια τέτοια ελπίδα δίνεται από το γεγονός ότι μετά τη δημοσίευση των εργασιών σχετικά με την πρόχειρη αλληλουχία του γονιδιώματος του ρυζιού, ο αριθμός των δημοσιεύσεων σχετικά με τη δομή αυτού του γονιδιώματος αυξήθηκε απότομα, ειδικότερα, εμφανίστηκαν πληροφορίες σχετικά με τη λεπτομερή αλληλουχία των χρωμοσωμάτων του 1 και 4.
Η γνώση, τουλάχιστον κατά προσέγγιση, του αριθμού των γονιδίων στα φυτά είναι θεμελιώδους σημασίας για τη συγκριτική γονιδιωματική των φυτών. Αρχικά πιστευόταν ότι αφού όλα τα ανθοφόρα φυτά είναι πολύ κοντά το ένα στο άλλο ως προς τα φαινοτυπικά τους χαρακτηριστικά, τα γονιδιώματά τους θα πρέπει επίσης να είναι κοντά. Και αν μελετήσουμε το γονιδίωμα Arabidopsis, θα λάβουμε πληροφορίες για τα περισσότερα γονιδιώματα άλλων φυτών. Έμμεση επιβεβαίωση αυτής της υπόθεσης παρέχεται από τα αποτελέσματα της αλληλούχισης του γονιδιώματος του ποντικιού, το οποίο είναι εκπληκτικά κοντά στο ανθρώπινο γονιδίωμα (περίπου 30 χιλιάδες γονίδια, από τα οποία μόνο 1.000 αποδείχθηκε ότι ήταν διαφορετικά).
Μπορεί να υποτεθεί ότι ο λόγος για τις διαφορές στα γονιδιώματα του Arabidopsis και του ρυζιού έγκειται στο ότι ανήκουν σε διαφορετικές κατηγορίες φυτών - δικοτυλήδονα και μονοκοτυλήδονα. Για να διευκρινιστεί αυτό το ζήτημα, είναι εξαιρετικά επιθυμητό να γνωρίζουμε τουλάχιστον την πρόχειρη πρωτογενή δομή κάποιου άλλου μονοκοτυλήδονου φυτού. Ο πιο ρεαλιστικός υποψήφιος μπορεί να είναι το καλαμπόκι, του οποίου το γονιδίωμα είναι περίπου ίσο με το ανθρώπινο γονιδίωμα, αλλά εξακολουθεί να είναι σημαντικά μικρότερο από τα γονιδιώματα άλλων δημητριακών. Η διατροφική αξία του καλαμποκιού είναι γνωστή.
Το τεράστιο υλικό που λαμβάνεται από τον προσδιορισμό της αλληλουχίας των γονιδιωμάτων του Arabidopsis και του ρυζιού γίνεται σταδιακά η βάση για μια μεγάλης κλίμακας μελέτη γονιδιωμάτων φυτών χρησιμοποιώντας μεθόδους συγκριτικής γονιδιωματικής. Τέτοιες μελέτες έχουν γενική βιολογική σημασία, καθώς καθιστούν δυνατό τον καθορισμό των βασικών αρχών της οργάνωσης του φυτικού γονιδιώματος στο σύνολό του και των επιμέρους χρωμοσωμάτων τους, τον εντοπισμό κοινών χαρακτηριστικών της δομής των γονιδίων και των ρυθμιστικών περιοχών τους και την εξέταση των σχέση μεταξύ του λειτουργικά ενεργού (γονιδίου) τμήματος του χρωμοσώματος και διαφόρων μη κωδικοποιών πρωτεϊνικών διαγονιδιακών περιοχών DNA. Η συγκριτική γενετική γίνεται επίσης ολοένα και πιο σημαντική για την ανάπτυξη της ανθρώπινης λειτουργικής γονιδιωματικής. Για συγκριτικές μελέτες αναλύθηκε η αλληλουχία των γονιδιωμάτων των ψαριών και των ποντικών.
Δεν είναι λιγότερο σημαντική η μελέτη μεμονωμένων γονιδίων που είναι υπεύθυνα για τη σύνθεση μεμονωμένων πρωτεϊνών που καθορίζουν συγκεκριμένες λειτουργίες του σώματος. Η πρακτική, κυρίως ιατρική, σημασία του προγράμματος Ανθρώπινου Γονιδιώματος έγκειται στην ανίχνευση, την απομόνωση, την αλληλούχιση και την καθιέρωση της λειτουργίας μεμονωμένων γονιδίων. Αυτή η περίσταση είχε επισημανθεί πριν από αρκετά χρόνια από τον J. Watson, ο οποίος τόνισε ότι το πρόγραμμα του ανθρώπινου γονιδιώματος θα ολοκληρωθεί μόνο όταν καθοριστούν οι λειτουργίες όλων των ανθρώπινων γονιδίων.
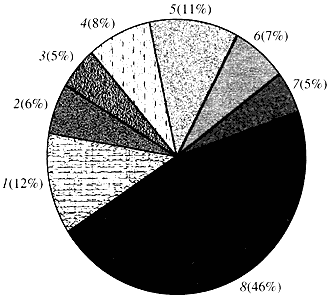
Ρύζι. 4.Ταξινόμηση ανά λειτουργία των γονιδίων Arabidopsis
1 - γονίδια για ανάπτυξη, διαίρεση και σύνθεση DNA. 2 - Γονίδια σύνθεσης RNA (μεταγραφή). 3 - γονίδια για σύνθεση και τροποποίηση πρωτεϊνών. 4 - γονίδια για ανάπτυξη, γήρανση και κυτταρικό θάνατο. 5 - γονίδια του κυτταρικού μεταβολισμού και του ενεργειακού μεταβολισμού. 6 - γονίδια για διακυτταρική αλληλεπίδραση και μετάδοση σήματος. 7 - γονίδια για την υποστήριξη άλλων κυτταρικών διεργασιών. 8 - γονίδια με άγνωστη λειτουργίαΌταν πρόκειται για τη λειτουργία των γονιδίων των φυτών, γνωρίζουμε λιγότερο από το ένα δέκατο αυτών που γνωρίζουμε για τα ανθρώπινα γονίδια. Ακόμη και στο Arabidopsis, του οποίου το γονιδίωμα είναι πολύ πιο μελετημένο από το ανθρώπινο γονιδίωμα, η λειτουργία σχεδόν των μισών γονιδίων του παραμένει άγνωστη (Εικ. 4). Εν τω μεταξύ, τα φυτά, εκτός από τα κοινά γονίδια στα ζώα, έχουν σημαντικό αριθμό γονιδίων ειδικά (ή τουλάχιστον κυρίως) σε αυτά. Μιλάμε για γονίδια που εμπλέκονται στη μεταφορά του νερού και στη σύνθεση κυτταρικών τοιχωμάτων, τα οποία απουσιάζουν στα ζώα, για γονίδια που διασφαλίζουν το σχηματισμό και τη λειτουργία των χλωροπλαστών, τη φωτοσύνθεση, τη δέσμευση αζώτου και τη σύνθεση πολυάριθμων αρωματικών προϊόντων. Αυτή η λίστα μπορεί να συνεχιστεί, αλλά είναι ήδη σαφές πόσο δύσκολο είναι το έργο που αντιμετωπίζει η λειτουργική γονιδιωματική των φυτών.
Η πλήρης αλληλουχία γονιδιώματος παρέχει σχεδόν αληθινές πληροφορίες για τον συνολικό αριθμό γονιδίων ενός δεδομένου οργανισμού, επιτρέπει περισσότερο ή λιγότερο λεπτομερείς και αξιόπιστες πληροφορίες για τη δομή τους να τοποθετούνται σε τράπεζες δεδομένων και διευκολύνει το έργο της απομόνωσης και της μελέτης μεμονωμένων γονιδίων. Ωστόσο, η αλληλουχία του γονιδιώματος δεν σημαίνει την καθιέρωση της λειτουργίας όλων των γονιδίων.
Μία από τις πιο πολλά υποσχόμενες προσεγγίσεις της λειτουργικής γονιδιωματικής βασίζεται στον εντοπισμό γονιδίων εργασίας στα οποία λαμβάνει χώρα η μεταγραφή του mRNA (ανάγνωση). Αυτή η προσέγγιση, συμπεριλαμβανομένης της χρήσης σύγχρονης τεχνολογίας μικροσυστοιχιών, καθιστά δυνατή την ταυτόχρονη ταυτοποίηση έως και δεκάδων χιλιάδων λειτουργικών γονιδίων. Πρόσφατα, χρησιμοποιώντας αυτή την προσέγγιση, ξεκίνησε η μελέτη των γονιδιωμάτων των φυτών. Για το Arabidopsis, ήταν δυνατό να ληφθούν περίπου 26 χιλιάδες μεμονωμένα μεταγραφήματα, γεγονός που διευκολύνει σημαντικά τη δυνατότητα προσδιορισμού της λειτουργίας σχεδόν όλων των γονιδίων του. Στις πατάτες, ήταν δυνατό να εντοπιστούν περίπου 20.000 χιλιάδες ενεργά γονίδια που είναι σημαντικά για την κατανόηση τόσο των διαδικασιών ανάπτυξης και σχηματισμού κονδύλων όσο και των διεργασιών της νόσου της πατάτας. Αναμένεται ότι αυτή η γνώση θα βελτιώσει την αντοχή ενός από τα σημαντικότερα τρόφιμα σε παθογόνους παράγοντες.
Μια λογική εξέλιξη της λειτουργικής γονιδιωματικής είναι η πρωτεομική. Αυτό το νέο πεδίο της επιστήμης μελετά το πρωτείωμα, το οποίο συνήθως αναφέρεται στο πλήρες σύνολο πρωτεϊνών σε ένα κύτταρο σε μια δεδομένη στιγμή. Αυτό το σύνολο πρωτεϊνών, που αντικατοπτρίζει τη λειτουργική κατάσταση του γονιδιώματος, αλλάζει συνεχώς, ενώ το γονιδίωμα παραμένει αμετάβλητο.
Η μελέτη των πρωτεϊνών έχει χρησιμοποιηθεί από καιρό για να γίνουν κρίσεις σχετικά με τη δραστηριότητα των γονιδιωμάτων των φυτών. Όπως είναι γνωστό, τα ένζυμα που βρίσκονται σε όλα τα φυτά διαφέρουν ως προς την αλληλουχία των αμινοξέων σε μεμονωμένα είδη και ποικιλίες. Τέτοια ένζυμα, με την ίδια λειτουργία, αλλά διαφορετικές αλληλουχίες μεμονωμένων αμινοξέων, ονομάζονται ισοένζυμα. Έχουν διαφορετικές φυσικοχημικές και ανοσολογικές ιδιότητες (μοριακό βάρος, φορτίο), οι οποίες μπορούν να ανιχνευθούν με χρωματογραφία ή ηλεκτροφόρηση. Για πολλά χρόνια, αυτές οι μέθοδοι έχουν χρησιμοποιηθεί με επιτυχία για τη μελέτη του λεγόμενου γενετικού πολυμορφισμού, δηλαδή των διαφορών μεταξύ οργανισμών, ποικιλιών, πληθυσμών, ειδών, ιδιαίτερα του σίτου και των σχετικών μορφών δημητριακών. Ωστόσο, πρόσφατα, λόγω της ταχείας ανάπτυξης των μεθόδων ανάλυσης DNA, συμπεριλαμβανομένης της αλληλουχίας, η μελέτη του πολυμορφισμού των πρωτεϊνών έχει αντικατασταθεί από τη μελέτη του πολυμορφισμού του DNA. Ωστόσο, η άμεση μελέτη των φασμάτων των πρωτεϊνών αποθήκευσης (προλαμίνες, γλιαδίνες κ.λπ.), που καθορίζουν τις βασικές θρεπτικές ιδιότητες των δημητριακών, παραμένει μια σημαντική και αξιόπιστη μέθοδος γενετικής ανάλυσης, επιλογής και παραγωγής σπόρων γεωργικών φυτών.
Η γνώση των γονιδίων, των μηχανισμών έκφρασης και ρύθμισής τους είναι εξαιρετικά σημαντική για την ανάπτυξη της βιοτεχνολογίας και την παραγωγή διαγονιδιακών φυτών. Είναι γνωστό ότι οι εντυπωσιακές επιτυχίες στον τομέα αυτό προκαλούν μικτές αντιδράσεις από την περιβαλλοντική και ιατρική κοινότητα. Ωστόσο, υπάρχει ένας τομέας της φυτικής βιοτεχνολογίας όπου αυτοί οι φόβοι, αν όχι εντελώς αβάσιμοι, τότε, σε κάθε περίπτωση, φαίνονται ασήμαντοι. Μιλάμε για δημιουργία διαγονιδιακών βιομηχανικών φυτών που δεν χρησιμοποιούνται ως προϊόντα διατροφής. Η Ινδία συγκέντρωσε πρόσφατα την πρώτη της καλλιέργεια διαγονιδιακού βαμβακιού που είναι ανθεκτικό σε μια σειρά από ασθένειες. Υπάρχουν πληροφορίες σχετικά με την εισαγωγή ειδικών γονιδίων που κωδικοποιούν πρωτεΐνες χρωστικής στο γονιδίωμα του βαμβακιού και την παραγωγή ινών βαμβακιού που δεν απαιτούν τεχνητή βαφή. Μια άλλη βιομηχανική καλλιέργεια που μπορεί να υποβληθεί σε αποτελεσματική γενετική μηχανική είναι το λινάρι. Η χρήση του ως εναλλακτική λύση του βαμβακιού για κλωστοϋφαντουργικές πρώτες ύλες έχει συζητηθεί πρόσφατα. Το πρόβλημα αυτό είναι εξαιρετικά σημαντικό για τη χώρα μας, η οποία έχει χάσει τις δικές της πηγές πρώτων υλών βαμβακιού.
ΠΡΟΟΠΤΙΚΕΣ ΓΙΑ ΤΗ ΜΕΛΕΤΗ ΤΩΝ ΦΥΤΩΝ ΓΟΝΙΔΩΜΑΤΩΝ
Είναι προφανές ότι οι δομικές μελέτες των φυτικών γονιδιωμάτων θα βασίζονται σε προσεγγίσεις και μεθόδους συγκριτικής γονιδιωματικής χρησιμοποιώντας ως κύριο υλικό τα αποτελέσματα της αποκρυπτογράφησης των γονιδιωμάτων του Arabidopsis και του ρυζιού. Σημαντικό ρόλο στην ανάπτυξη της συγκριτικής γονιδιωματικής των φυτών θα παίξει, χωρίς αμφιβολία, η πληροφορία που αργά ή γρήγορα θα παράσχει η συνολική (πρόχειρη) αλληλουχία των γονιδιωμάτων άλλων φυτών. Σε αυτή την περίπτωση, η συγκριτική γονιδιωματική των φυτών θα βασίζεται στη δημιουργία γενετικών σχέσεων μεταξύ μεμονωμένων τόπων και χρωμοσωμάτων που ανήκουν σε διαφορετικά γονιδιώματα. Θα μιλήσουμε όχι τόσο για τη γενική γονιδιωματική των φυτών, αλλά για την επιλεκτική γονιδιωματική των μεμονωμένων χρωμοσωμικών τόπων. Έτσι, πρόσφατα αποδείχθηκε ότι το γονίδιο που είναι υπεύθυνο για την εαρινοποίηση βρίσκεται στον τόπο VRn-AI του χρωμοσώματος 5Α του εξαπλοειδούς σίτου και στον τόπο Hd-6 του χρωμοσώματος 3 του ρυζιού.
Η ανάπτυξη αυτών των μελετών θα αποτελέσει μια ισχυρή ώθηση για την ταυτοποίηση, την απομόνωση και τον προσδιορισμό της αλληλουχίας πολλών λειτουργικά σημαντικών γονιδίων φυτών, ιδιαίτερα των γονιδίων που είναι υπεύθυνα για την αντοχή στις ασθένειες, την αντοχή στην ξηρασία και την προσαρμοστικότητα σε διάφορες συνθήκες ανάπτυξης. Η λειτουργική γονιδιωματική, που βασίζεται στη μαζική ταυτοποίηση (διαλογή) των γονιδίων που λειτουργούν στα φυτά, θα χρησιμοποιείται όλο και περισσότερο.
Μπορούμε να προβλέψουμε περαιτέρω βελτιώσεις στις χρωμοσωμικές τεχνολογίες, κυρίως στη μέθοδο μικροδιατομής. Η χρήση του διευρύνει δραματικά τις δυνατότητες γονιδιωματικής έρευνας χωρίς να απαιτεί τεράστιο κόστος, όπως η ολική αλληλουχία γονιδιώματος. Η μέθοδος εντοπισμού μεμονωμένων γονιδίων σε φυτικά χρωμοσώματα με χρήση υβριδισμού θα γίνει πιο διαδεδομένη. επί τόπου.Προς το παρόν, η χρήση του περιορίζεται από τον τεράστιο αριθμό επαναλαμβανόμενων αλληλουχιών στο φυτικό γονιδίωμα και πιθανώς από τις ιδιαιτερότητες της δομικής οργάνωσης των φυτικών χρωμοσωμάτων.
Στο άμεσο μέλλον, οι χρωμοσωμικές τεχνολογίες θα αποκτήσουν επίσης μεγάλη σημασία για την εξελικτική γονιδιωματική των φυτών. Αυτές οι τεχνολογίες, οι οποίες είναι σχετικά φθηνές, καθιστούν δυνατή την ταχεία αξιολόγηση της ενδοειδικής και ενδοειδικής μεταβλητότητας και τη μελέτη πολύπλοκων αλλοπολυπλοειδών γονιδιωμάτων τετραπλοειδούς και εξαπλοειδούς σίτου και τριτικάλε. να αναλύσει τις εξελικτικές διαδικασίες σε χρωμοσωμικό επίπεδο. Διερεύνηση του σχηματισμού συνθετικών γονιδιωμάτων και της εισαγωγής (εισόδου) ξένου γενετικού υλικού. προσδιορίζει τις γενετικές σχέσεις μεταξύ μεμονωμένων χρωμοσωμάτων διαφορετικών ειδών.
Για τον χαρακτηρισμό του γονιδιώματος θα χρησιμοποιηθεί η μελέτη του καρυότυπου του φυτού με χρήση κλασικών κυτταρογενετικών μεθόδων, εμπλουτισμένων με μοριακή βιολογική ανάλυση και τεχνολογίες υπολογιστών. Αυτό είναι ιδιαίτερα σημαντικό για τη μελέτη της σταθερότητας και της μεταβλητότητας του καρυότυπου σε επίπεδο όχι μόνο μεμονωμένων οργανισμών, αλλά και πληθυσμών, ποικιλιών και ειδών. Τέλος, είναι δύσκολο να φανταστεί κανείς πώς μπορεί κανείς να εκτιμήσει τον αριθμό και τα φάσματα των χρωμοσωμικών αναδιατάξεων (εκτροπές, γέφυρες) χωρίς τη χρήση μεθόδων διαφορικής χρώσης. Τέτοιες μελέτες είναι εξαιρετικά ελπιδοφόρες για την παρακολούθηση του περιβάλλοντος με βάση την κατάσταση του φυτικού γονιδιώματος.
Στη σύγχρονη Ρωσία, είναι απίθανο να πραγματοποιηθεί απευθείας αλληλούχιση των γονιδιωμάτων των φυτών. Τέτοιες εργασίες, που απαιτούν μεγάλες επενδύσεις, είναι μη βιώσιμες για τη σημερινή μας οικονομία. Εν τω μεταξύ, πληροφορίες σχετικά με τη δομή των γονιδιωμάτων του Arabidopsis και του ρυζιού, που λαμβάνονται από την παγκόσμια επιστήμη και είναι διαθέσιμες σε διεθνείς τράπεζες δεδομένων, είναι επαρκείς για την ανάπτυξη της γονιδιωματικής των εγχώριων φυτών. Είναι δυνατό να προβλεφθεί επέκταση της έρευνας σε γονιδιώματα φυτών με βάση προσεγγίσεις συγκριτικής γονιδιωματικής για την επίλυση συγκεκριμένων προβλημάτων αναπαραγωγής και φυτικής παραγωγής, καθώς και για τη μελέτη της προέλευσης διαφόρων ειδών φυτών οικονομικής σημασίας.
Μπορούμε να υποθέσουμε ότι στην οικιακή πρακτική αναπαραγωγής και στην καλλιέργεια φυτών, θα χρησιμοποιηθούν ευρέως γονιδιωματικές προσεγγίσεις όπως ο γενετικός τύπος (αναλύσεις RELF, RAPD, AFLP, κ.λπ.), οι οποίες είναι αρκετά προσιτές για τον προϋπολογισμό μας. Παράλληλα με τις άμεσες μεθόδους προσδιορισμού του πολυμορφισμού του DNA, προσεγγίσεις που βασίζονται στη μελέτη του πολυμορφισμού των πρωτεϊνών, κυρίως πρωτεϊνών αποθήκευσης δημητριακών, θα χρησιμοποιηθούν για την επίλυση προβλημάτων γενετικής και αναπαραγωγής φυτών. Οι τεχνολογίες χρωμοσωμάτων θα χρησιμοποιηθούν ευρέως. Είναι σχετικά φθηνά και η ανάπτυξή τους απαιτεί αρκετά μέτριες επενδύσεις. Στον τομέα της έρευνας των χρωμοσωμάτων, η εγχώρια επιστήμη δεν είναι κατώτερη από τον κόσμο.
Πρέπει να τονιστεί ότι η επιστήμη μας έχει συμβάλει σημαντικά στη διαμόρφωση και ανάπτυξη της γονιδιωματικής των φυτών [,].
Τον θεμελιώδη ρόλο έπαιξε ο Ν.Ι. Βαβίλοφ (1887-1943).
Στη μοριακή βιολογία και τη γονιδιωματική των φυτών είναι εμφανής η πρωτοποριακή συμβολή του Α.Ν. Belozersky (1905-1972).
Στον τομέα της έρευνας των χρωμοσωμάτων, είναι απαραίτητο να σημειωθεί το έργο του εξαιρετικού γενετιστή S.G. Navashin (1857-1930), ο οποίος ανακάλυψε για πρώτη φορά δορυφορικά χρωμοσώματα στα φυτά και απέδειξε ότι είναι δυνατό να διακρίνουμε μεμονωμένα χρωμοσώματα από τα χαρακτηριστικά της μορφολογίας τους.
Ένα άλλο κλασικό της ρωσικής επιστήμης G.A. Ο Levitsky (1878-1942) περιέγραψε λεπτομερώς τα χρωμοσώματα της σίκαλης, του σιταριού, του κριθαριού, του μπιζελιού και των ζαχαρότευτλων, εισήγαγε τον όρο «καρυότυπος» στην επιστήμη και ανέπτυξε το δόγμα του.
Οι σύγχρονοι ειδικοί, βασιζόμενοι στα επιτεύγματα της παγκόσμιας επιστήμης, μπορούν να συμβάλουν σημαντικά στην περαιτέρω ανάπτυξη της φυτικής γενετικής και γονιδιωματικής.
Ο συγγραφέας εκφράζει την εγκάρδια ευγνωμοσύνη του στον Ακαδημαϊκό Yu.P. Altukhov για κριτική συζήτηση του άρθρου και πολύτιμες συμβουλές.Το έργο της ομάδας με επικεφαλής τον συγγραφέα του άρθρου υποστηρίχθηκε από το Ρωσικό Ίδρυμα Βασικής Έρευνας (επιχορηγήσεις Αρ. 99-04-48832; 00-04-49036; 00-04-81086), το Πρόγραμμα του Προέδρου του τη Ρωσική Ομοσπονδία για την υποστήριξη επιστημονικών σχολών (επιχορηγήσεις αρ. 00-115 -97833 και NSh-1794.2003.4) και το Πρόγραμμα της Ρωσικής Ακαδημίας Επιστημών «Μοριακοί γενετικοί και χρωμοσωμικοί δείκτες στην ανάπτυξη σύγχρονων μεθόδων επιλογής και σποράς παραγωγή."
ΒΙΒΛΙΟΓΡΑΦΙΑ
1. Zelenin A.V., Badaeva E.D., Muravenko O.V.Εισαγωγή στη γονιδιωματική των φυτών // Μοριακή βιολογία. 2001. Τ. 35. σ. 339-348. 2. Στυλό Ε. Bonanza for Plant Genomics // Science. 1998. V. 282. Σ. 652-654. 3. Γονιδιωματική των φυτών // Proc. Natl. Ακαδ. Sci. ΗΠΑ. 1998. V. 95. Σ. 1962-2032. 4. Kartel N.A. και τα λοιπά.Γενεσιολογία. Εγκυκλοπαιδικό Λεξικό. Μινσκ: Technologia, 1999. 5. Badaeva E.D., Friebe B., Gill B.S. 1996. Διαφοροποίηση γονιδιώματος στον Αιγίλωψ. 1. Κατανομή άκρως επαναλαμβανόμενων αλληλουχιών DNA σε χρωμοσώματα διπλοειδών ειδών // Γονιδίωμα. 1996. V. 39. Σ. 293-306. History of chromosome analysis // Biol. μεμβράνες. 2001. Τ. 18. σ. 164-172.
Για δύο από τα πιο έγκυρα επιστημονικά περιοδικά του κόσμου - το British Nature και το American Science - είναι εξαιρετικά σπάνιο να αφιερώνουν ταυτόχρονα ένα σημαντικό μέρος των επόμενων τευχών τους στο ίδιο θέμα. Και αν συμβεί, δείχνει την εξαιρετική σημασία αυτού του θέματος. Έτσι, η δημοσίευση 12 άρθρων ταυτόχρονα αφιερωμένων στην αποκωδικοποίηση του γονιδιώματος του χιμπατζή και τη σύγκρισή του με το ανθρώπινο γονιδίωμα είναι, φυσικά, ένα εξαιρετικό γεγονός.
Μια διεθνής κοινοπραξία δημιουργήθηκε για την υλοποίηση ενός έργου για τη χαρτογράφηση και τη συγκριτική ανάλυση του γονιδιώματος του χιμπατζή. Συμμετείχαν 67 επιστήμονες από 23 επιστημονικά ιδρύματα σε 5 χώρες - ΗΠΑ, Ισραήλ, Ισπανία, Ιταλία και Γερμανία. Η εργασία συντονίστηκε από γενετιστές από το Πανεπιστήμιο του Χάρβαρντ και το Ινστιτούτο Τεχνολογίας της Μασαχουσέτης στη Βοστώνη. Και αίμα για ανάλυση DNA δόθηκε από έναν νεαρό αρσενικό χιμπατζή ονόματι Κλιντ, κάτοικος ενός από τους καταλύτες του Εθνικού Κέντρου Ερευνών Πρωτευόντων Yerkes στην Ατλάντα της Τζόρτζια. Δυστυχώς, τον Ιανουάριο του τρέχοντος έτους ο δότης πέθανε από οξεία καρδιακή ανεπάρκεια στην ακμή της ζωής του, σε ηλικία 24 ετών. Ο σκελετός του εκτίθεται τώρα στο Field Museum στο Σικάγο. Ωστόσο, η πιο σημαντική αξία που κληρονόμησε η ανθρωπότητα από τον Κλιντ είναι ένα μέρος του αίματός του, το οποίο χρησίμευσε ως το αρχικό υλικό για την αποκρυπτογράφηση και την ανάλυση του γονιδιώματος του χιμπατζή. Τώρα τα πρωτεύοντα έχουν ενταχθεί στον κατάλογο των οργανισμών των οποίων το γενετικό υλικό έχει χαρτογραφηθεί πλήρως. Αυτή η λίστα σήμερα περιλαμβάνει ήδη εκατοντάδες είδη: υπάρχουν μούχλες, βακτήρια, συμπεριλαμβανομένων των αιτιολογικών παραγόντων επικίνδυνων μολυσματικών ασθενειών (άνθρακας, τουλαραιμίας, πανώλης, τύφος) και φυτά (ρύζι, καφεόδεντρο) και έντομα (κουνούπι ελονοσίας) και πτηνά (π.χ. κοτόπουλο) και θηλαστικά (ποντίκι, αρουραίος, σκύλος, γουρούνι, αγελάδα). Ωστόσο, οι ανθρωποειδείς πίθηκοι καταλαμβάνουν, φυσικά, μια πολύ ιδιαίτερη θέση σε αυτή τη λίστα. Σύμφωνα με τον Robert Waterston, διευθυντή γονιδιωματικής έρευνας στο University of Washington Graduate School of Medicine στο Σιάτλ, «η μελέτη των χιμπατζήδων, ως ο πλησιέστερος ζωντανός συγγενής στους ανθρώπους στη Γη, μπορεί να μας δώσει τις περισσότερες πληροφορίες για τον εαυτό μας». Ωστόσο, προτού προχωρήσω στη συζήτηση των αποτελεσμάτων που προέκυψαν από τους επιστήμονες, θα επιτρέψω στον εαυτό μου μια μικρή παρέκβαση -ή, αν θέλετε, μια υπενθύμιση- για να γίνει πιο σαφές για τι πράγμα μιλάμε.
Όπως γνωρίζετε, κάθε ζωντανός οργανισμός αποτελείται από κύτταρα και στον πυρήνα κάθε κυττάρου υπάρχει το ίδιο σύνολο γενετικών πληροφοριών χαρακτηριστικών ενός δεδομένου βιολογικού είδους. Αυτό το σύνολο ονομάζεται γονιδίωμα. Τα χρωμοσώματα είναι ο φορέας της γενετικής πληροφορίας. Ένα χρωμόσωμα είναι ένα μόριο δεοξυριβονουκλεϊκού οξέος (DNA για συντομία) και αποτελείται από δύο μακριές πολυνουκλεοτιδικές αλυσίδες στριμμένες η μία γύρω από την άλλη και συνδεδεμένες μεταξύ τους με τους λεγόμενους δεσμούς υδρογόνου. Αυτό το μόριο ονομάζεται διπλή έλικα και μπορεί κάπως απλοποιημένα να φανταστεί ως μια σκάλα με στριφτό σχοινί. Διαφορετικά είδη ζώων έχουν διαφορετικό αριθμό χρωμοσωμάτων. Έτσι, το ανθρώπινο γονιδίωμα αποτελείται από 23 ζεύγη χρωμοσωμάτων - σε κάθε ζευγάρι, το ένα χρωμόσωμα προέρχεται από τον πατέρα και το άλλο από τη μητέρα. Η μύγα - Drosophila - περιέχει 4 ζεύγη χρωμοσωμάτων στους πυρήνες των κυττάρων της, ενώ, για παράδειγμα, τα βακτήρια έχουν μόνο ένα ασύζευκτο χρωμόσωμα. Τα γονίδια βρίσκονται στα χρωμοσώματα σε αυστηρά καθορισμένες περιοχές - ένα είδος μονάδας κληρονομικότητας. Χημικά, τα γονίδια αποτελούνται από μόρια 4 αζωτούχων ενώσεων - αδενίνη, κυτοσίνη, γουανίνη και θυμίνη. Αυτές οι αποκαλούμενες βάσεις νουκλεοτιδίων επαναλαμβάνονται με αυστηρά καθορισμένη σειρά, σχηματίζοντας ζεύγη αδενίνης-θυμίνης και γουανίνης-κυτοσίνης. Ένα μόνο γονίδιο μπορεί να περιέχει από πολλές χιλιάδες έως περισσότερες από δύο εκατομμύρια βάσεις νουκλεοτιδίων. Είναι η αλληλουχία τους που καθορίζει τις συγκεκριμένες λειτουργίες κάθε συγκεκριμένου γονιδίου.
Μεταφορικά, το γονιδίωμα μπορεί να φανταστεί ως εξής: ο κυτταρικός πυρήνας είναι μια βιβλιοθήκη στην οποία αποθηκεύονται οδηγίες για τη διασφάλιση της ζωής. Τα χρωμοσώματα παίζουν το ρόλο των ραφιών. υπάρχουν βιβλία στα ράφια - μόρια DNA. τα γονίδια είναι κεφάλαια μέσα στα βιβλία και οι βάσεις νουκλεοτιδίων - αδενίνη, θυμίνη, γουανίνη και κυτοσίνη, που συνήθως υποδηλώνονται με τα αρχικά γράμματα των ονομάτων τους A, T, G και C - αυτό είναι το ίδιο το αλφάβητο με το οποίο είναι το κείμενο του γονιδιώματος γραπτός. Το ανθρώπινο γονιδίωμα, για παράδειγμα, είναι μια αλυσίδα 3 δισεκατομμυρίων 200 εκατομμυρίων γραμμάτων.
Αλλά το γεγονός ότι τα γονίδια υπάρχουν και ότι λειτουργούν δεν αρκεί: πρέπει να λειτουργούν με διαφορετικούς τρόπους, παρέχοντας ορισμένες συγκεκριμένες λειτουργίες. Άλλωστε, τα κύτταρα διαφορετικών οργάνων και ιστών - ας πούμε, του δέρματος, του ήπατος, της καρδιάς και του εγκεφάλου - είναι εντυπωσιακά διαφορετικά μεταξύ τους. Εν τω μεταξύ, ο πυρήνας καθενός από αυτούς περιέχει το ίδιο σύνολο γονιδίων. Είναι όλα σχετικά με τη γονιδιακή δραστηριότητα: ορισμένα γονίδια λειτουργούν σε ορισμένα κύτταρα και άλλα λειτουργούν σε άλλα. Τα χρωμοσώματα λοιπόν είναι φορείς όχι μόνο γονιδίων, αλλά και εκείνων των πρωτεϊνικών παραγόντων που ελέγχουν τις λειτουργίες τους. Αυτό το σύνολο γονιδίων, μαζί με ρυθμιστικά στοιχεία, αποτελεί τη δομή μέσα στο κύτταρο που παρέχει όλες τις απαραίτητες λειτουργίες.
Και τώρα, οπλισμένοι με αυτή τη γνώση, ας επιστρέψουμε στα αποτελέσματα που προέκυψαν κατά την αποκρυπτογράφηση του γονιδιώματος του χιμπατζή. Για προφανείς λόγους, το μεγαλύτερο ενδιαφέρον τόσο μεταξύ των ειδικών όσο και του ευρύτερου κοινού είναι ο κατάλογος εκείνων των διαφορών στους γενετικούς κώδικες των χιμπατζήδων και των ανθρώπων που έχουν συσσωρευτεί τα τελευταία 6 εκατομμύρια χρόνια, από την εξελικτική πορεία δύο ειδών που είχαν κοινό πρόγονο . σε διασταση. Ο Svante Pääbo, ερευνητής στο Ινστιτούτο Max Planck για την Εξελικτική Ανθρωπολογία στη Λειψία και ένας από τους συμμετέχοντες στο έργο, αξιολογεί τη βάση δεδομένων που προκύπτει ως εξής:
Είναι ένα εξαιρετικά χρήσιμο εργαλείο που θα μας βοηθήσει να βρούμε μια απάντηση στο ερώτημα ποιες γενετικές μεταλλάξεις εξηγούν την εντυπωσιακή διαφορά μεταξύ του ανθρώπου ως βιολογικού είδους και όλων των άλλων ζωικών ειδών. Μία από τις κατευθύνσεις αυτής της αναζήτησης καταλήγει στην προσπάθεια εντοπισμού της σχέσης μεταξύ γενετικών διαφορών και της δραστηριότητας ορισμένων γονιδίων.
Πρώτα απ 'όλα, πρέπει να σημειωθεί ότι τα δεδομένα που ελήφθησαν εξέπληξαν τους ειδικούς. Η κύρια έκπληξη είναι ότι το γονιδίωμα του χιμπατζή αποδείχθηκε ότι ήταν 98,8 τοις εκατό πανομοιότυπο με το ανθρώπινο γονιδίωμα. Σε γενικές γραμμές, η γενετική ομοιότητα μεταξύ ανθρώπων και χιμπατζήδων είναι 10 φορές μεγαλύτερη από ό,τι μεταξύ ποντικών και αρουραίων. Οι ερασιτέχνες πιθανότατα θα εντυπωσιαστούν από μια τόσο μεγάλη ομοιότητα, αυτή την σχεδόν πλήρη ταυτότητα γονιδιωμάτων, αλλά οι επιστήμονες εξεπλάγησαν από το ακριβώς αντίθετο: το γεγονός ότι η διαφορά αποδείχθηκε αρκετά σημαντική. Επιπλέον, αυτό το ποσοστό - σύμπτωση 98,8% - δεν αντικατοπτρίζει πλήρως την κατάσταση των πραγμάτων. Λαμβάνεται συγκρίνοντας μεμονωμένα γράμματα του γενετικού κώδικα στο κωδικοποιητικό DNA. Εδώ, οι επιστήμονες μέτρησαν 35 εκατομμύρια αποκλίσεις, οι οποίες ανήλθαν στο 1,2 τοις εκατό ολόκληρου του γονιδιώματος του χιμπατζή, το οποίο έχει περίπου 3 δισεκατομμύρια 100 εκατομμύρια ζεύγη νουκλεοτιδίων. Αλλά δεν είναι μόνο αυτό: ανακαλύφθηκαν επίσης σημαντικές διαφορές στην κατανομή αυτών των αλληλουχιών νουκλεοτιδικών βάσεων που σχηματίζουν μη κωδικοποιητικό, «εγωιστικό» DNA. Αυτές οι αναντιστοιχίες αντιπροσώπευαν ένα άλλο 2,7 τοις εκατό του συνόλου του γονιδιώματος, σε σύνολο σχεδόν 4 τοις εκατό.
Συνολικά, οι χιμπατζήδες δεν είχαν 53 γονίδια που έχουν οι άνθρωποι. Συγκεκριμένα, από το γονιδίωμα του χιμπατζή λείπουν τρία γονίδια που παίζουν βασικό ρόλο στην ανάπτυξη φλεγμονής, η οποία είναι γνωστό ότι προκαλεί πολλές ανθρώπινες ασθένειες. Από την άλλη, οι άνθρωποι φαίνεται να έχουν χάσει στη διαδικασία της εξέλιξης ένα γονίδιο που προστατεύει τα ζώα από τη νόσο του Αλτσχάιμερ.
Οι πιο σημαντικές διαφορές αφορούν γονίδια που ρυθμίζουν το ανοσοποιητικό σύστημα. Σύμφωνα με τον καθηγητή Evan Eichler, συνεργάτη στο University of Washington Graduate School of Medicine στο Σιάτλ, αυτό δείχνει ότι οι χιμπατζήδες και οι άνθρωποι έπρεπε να αντιμετωπίσουν διαφορετικά παθογόνα και να καταπολεμήσουν διαφορετικές ασθένειες κατά την εξελικτική τους ανάπτυξη. Ο Svante Pääbo εξηγεί:
Πρώτα απ 'όλα, αναρωτηθήκαμε ποια τμήματα DNA θα μπορούσαν να παρέχουν πληροφορίες για την προέλευση ορισμένων ασθενειών. Γνωρίζουμε ότι ορισμένες από τις γενετικές δομές που προκαλούν μια συγκεκριμένη ασθένεια βρίσκονται τόσο στους χιμπατζήδες όσο και στους ανθρώπους. Προφανώς, αυτές οι δομές κληρονόμησαν και τα δύο είδη από τον κοινό τους πρόγονο. Ωστόσο, υπάρχουν ασθένειες για τις οποίες προέκυψε γενετική προδιάθεση κατά τη διαδικασία της εξέλιξης μόνο στους ανθρώπους. Σε αυτές τις περιπτώσεις, η συγκριτική ανάλυση DNA θα μας δώσει πολύτιμες πληροφορίες για τη γενετική φύση τέτοιων ασθενειών και την ευαισθησία του ανθρώπου ως είδους σε αυτές.
Αναλύοντας τα δεδομένα που συλλέχθηκαν, οι επιστήμονες έκαναν ένα είδος επικάλυψης υπολογιστή του χάρτη γονιδιώματος του χιμπατζή στον χάρτη του ανθρώπινου γονιδιώματος, που τους επέτρεψε να αναγνωρίσουν τρεις κατηγορίες λεγόμενων διπλασιασμών DNA - αυτές που υπάρχουν στο ανθρώπινο γονιδίωμα, αλλά απουσιάζουν στο το γονιδίωμα του χιμπατζή, αυτά που υπάρχουν στο γονιδίωμα των χιμπατζήδων, αλλά απουσιάζουν από το ανθρώπινο γονιδίωμα, και αυτά που υπάρχουν στο γονιδίωμα και των δύο ειδών. Ο διπλασιασμός του DNA είναι μια μορφή μετάλλαξης κατά την οποία ένα τμήμα ενός χρωμοσώματος διπλασιάζεται. Σε αυτή την περίπτωση, λήφθηκαν υπόψη τμήματα DNA με μήκος τουλάχιστον 20 χιλιάδων ζευγών νουκλεοτιδίων. Αποδείχθηκε ότι περίπου το ένα τρίτο των διπλασιασμών DNA που βρέθηκαν στους ανθρώπους απουσιάζουν στους χιμπατζήδες. Σύμφωνα με τον Eikler, ο αριθμός αυτός εξέπληξε τους γενετιστές γιατί υποδηλώνει πολύ υψηλή συχνότητα μεταλλάξεων σε σύντομο χρονικό διάστημα, σύμφωνα με τα εξελικτικά πρότυπα. Ταυτόχρονα, μια ανάλυση των διπλασιασμών DNA μοναδικών για το γονιδίωμα του χιμπατζή έδειξε ότι αν και ο αριθμός των τόπων όπου εμφανίζονται είναι σχετικά μικρός, ο αριθμός των αντιγράφων των διπλών τμημάτων είναι πολύ μεγαλύτερος από ό,τι στους ανθρώπους. Και σε περιπτώσεις όπου ο διπλασιασμός του DNA συμβαίνει τόσο σε χιμπατζήδες όσο και σε ανθρώπους, στους χιμπατζήδες συνήθως αντιπροσωπεύεται από μεγάλο αριθμό αντιγράφων. Συγκεκριμένα, οι επιστήμονες ανακάλυψαν ένα τμήμα που εμφανίζεται 4 φορές στο ανθρώπινο γονιδίωμα και 400 φορές στο γονιδίωμα του χιμπατζή. Είναι ενδιαφέρον ότι αυτή η περιοχή βρίσκεται κοντά στην περιοχή που στους χιμπατζήδες και άλλους μεγάλους πιθήκους χωρίζεται σε 2 χρωμοσώματα και στους ανθρώπους συντήκεται σε ένα - το χρωμόσωμα Νο. 2.
Ωστόσο, οι εντυπωσιακές διαφορές μεταξύ των πιθήκων και των ανθρώπων εξηγούνται όχι τόσο από διαφορές στον γενετικό κώδικα, αλλά από διαφορετικές γονιδιακές δραστηριότητες, τονίζει ο Svante Päbo. Μια ομάδα ερευνητών με επικεφαλής τον ίδιο μελέτησε και συνέκρινε τη δραστηριότητα 21 χιλιάδων γονιδίων στα κύτταρα της καρδιάς, του ήπατος, των νεφρών, των όρχεων και του εγκεφάλου και των δύο πρωτευόντων. Αποδείχθηκε ότι δεν υπάρχει πλήρης σύμπτωση γονιδιακής δραστηριότητας σε κανένα από αυτά τα όργανα, αλλά οι διαφορές κατανέμονται εξαιρετικά άνισα. Παραδόξως, οι επιστήμονες κατέγραψαν τις μικρότερες διαφορές στα εγκεφαλικά κύτταρα - ανήλθαν μόνο σε λίγα τοις εκατό. Και οι μεγαλύτερες διαφορές βρέθηκαν στους όρχεις: εδώ κάθε τρίτο γονίδιο έχει διαφορετική δραστηριότητα. Ωστόσο, αυτό είναι αρκετά κατανοητό αν λάβουμε υπόψη ότι οι χιμπατζήδες δεν σχηματίζουν μονογαμικές οικογένειες, αλλά ζουν σε ομάδες, ένα είδος κοινοτήτων, που αριθμούν 25-30 άτομα και των δύο φύλων. Δηλαδή, η «ασωτία» μεταξύ των χιμπατζήδων είναι πολύ πιο διαδεδομένη από ό,τι μεταξύ των ανθρώπων. Για να αυξήσουν τις πιθανότητές τους να τεκνοποιήσουν σε ακατάλληλες συνθήκες, οι αρσενικοί χιμπατζήδες πρέπει να παράγουν τεράστιες ποσότητες σπέρματος. Δεν είναι τυχαίο ότι οι όρχεις τους είναι δέκα φορές μεγαλύτεροι από αυτούς των ανδρών homo sapiens. Αλλά δεν είναι μόνο το μέγεθος, φυσικά, λέει ο Svante Päbo:
Τα δεδομένα μας υποδεικνύουν πολύ υψηλή δραστηριότητα εκείνων των γονιδίων στο χρωμόσωμα Υ που είναι άμεσα υπεύθυνα για την παραγωγή σπέρματος.
Και για το γεγονός ότι οι άνθρωποι είναι σωματικά πολύ πιο αδύναμοι από τους χιμπατζήδες, οι επιστήμονες βρήκαν μια γενετική εξήγηση: στους πιθήκους, οι μύες λειτουργούν 5-7 φορές πιο αποτελεσματικά επειδή σε όλους τους εκπροσώπους της ανθρώπινης φυλής, το γονίδιο MYH16, το οποίο κωδικοποιεί τη "μυοσίνη" - μια πρωτεΐνη μυϊκής ίνας - αντιπροσωπεύεται από ένα μεταλλαγμένο αντίγραφο.
Ωστόσο, εάν επικεντρωθούμε στο ερώτημα ποια είναι η κύρια γενετική διαφορά μεταξύ του ανθρώπου ως βιολογικού είδους και των πιθήκων και τι εξηγεί μια τόσο επιτυχημένη επέκταση των ανθρώπων κατά την εξέλιξη, τότε η απάντηση, προφανώς, θα πρέπει να αναζητηθεί στις 6 περιοχές του γονιδίωμα που προσδιορίστηκε από επιστήμονες. Στο ανθρώπινο γονιδίωμα, αυτές οι περιοχές, που περιέχουν συνολικά αρκετές εκατοντάδες γονίδια, είναι τόσο σταθερές που είναι πρακτικά ίδιες σε όλους τους ανθρώπους. στο γονιδίωμα του χιμπατζή, αντίθετα, συχνά περιέχουν μεταλλάξεις. Προφανώς, πιστεύουν οι επιστήμονες, αυτές οι περιοχές έπαιξαν εξαιρετικά σημαντικό ρόλο στη διαδικασία της εξέλιξής μας. Είναι αξιοσημείωτο ότι το γονίδιο FOXP2 βρίσκεται σε μία από αυτές τις περιοχές, ένα από τα 4 γονίδια που είναι υπεύθυνα για την ανάπτυξη της ομιλίας. Όπως έχουν δείξει πειράματα, σε εργαστηριακές συνθήκες οι πίθηκοι είναι σε θέση να μάθουν ένα αρκετά σημαντικό σύνολο σημείων και συμβόλων. Οι χιμπατζήδες που ζουν στη φύση χρησιμοποιούν μια πολύ πλούσια γκάμα ήχων για επικοινωνία. Ωστόσο, δεν μπορούν σωματικά να κάνουν τις κινήσεις με τα χείλη και τη γλώσσα τους που είναι απαραίτητες για την άρθρωση του λόγου. Ίσως ήταν η μετάλλαξη του γονιδίου FOXP2 που έγινε ένας από τους βασικούς παράγοντες που καθόρισε τόσο διαφορετικές εξελικτικές τύχες διαφορετικών ειδών πρωτευόντων.
Ωστόσο, δεν πρέπει να ξεχνάμε ότι ο άνθρωπος ξεχώριζε μεταξύ άλλων ζωικών ειδών όχι μόνο λόγω της ανεπτυγμένης ομιλίας του. Αλλά ποιες γενετικές δομές προκαθόρισαν την όρθια στάση και την ταχεία αύξηση του όγκου του εγκεφάλου, που συνεπαγόταν οτιδήποτε άλλο, είτε ήταν η δημιουργία εργαλείων είτε η χρήση φωτιάς - οι επιστήμονες δεν διακινδυνεύουν ακόμη καν να εκφράσουν υποθέσεις σχετικά με αυτό το σκορ.
πλήρως καθορισμένο. Επομένως, η εργασία για την αποκρυπτογράφηση του γονιδιώματος των νηματωδών θα πρέπει να θεωρείται πολύ επιτυχημένη.
Ακόμη μεγαλύτερη επιτυχία συνδέεται με την αποκρυπτογράφηση του γονιδιώματος της Drosophila, μόνο σε
2 φορές μικρότερο σε μέγεθος από το ανθρώπινο DNA και 20 φορές μεγαλύτερο από το DNA των νηματωδών. Παρά τον υψηλό βαθμό γενετικής γνώσης της Drosophila, περίπου το 10% των γονιδίων της ήταν άγνωστα μέχρι αυτή τη στιγμή. Αλλά το πιο παράδοξο είναι ότι το Drosophila, το οποίο είναι πολύ πιο οργανωμένο σε σύγκριση με το νηματώδη, έχει λιγότερα γονίδια από ένα μικροσκοπικό στρογγυλό σκουλήκι! Από σύγχρονη βιολογική άποψη, αυτό είναι δύσκολο να εξηγηθεί. Περισσότερα γονίδια από αυτά της Drosophila υπάρχουν επίσης στο αποκρυπτογραφημένο γονιδίωμα ενός φυτού από την οικογένεια των σταυρανθών - Arabidopsis, που χρησιμοποιείται ευρέως από τους γενετιστές ως κλασικό πειραματικό αντικείμενο.
Η ανάπτυξη γονιδιωματικών έργων συνοδεύτηκε από εντατική ανάπτυξη σε πολλούς τομείς της επιστήμης και της τεχνολογίας. Έτσι, η βιοπληροφορική έλαβε μια ισχυρή ώθηση για την ανάπτυξή της. Δημιουργήθηκε μια νέα μαθηματική συσκευή για την αποθήκευση και την επεξεργασία τεράστιων ποσοτήτων πληροφοριών. έχουν σχεδιαστεί συστήματα υπερυπολογιστών με πρωτοφανή ισχύ. Έχουν γραφτεί χιλιάδες προγράμματα που επιτρέπουν, μέσα σε λίγα λεπτά, τη διεξαγωγή συγκριτικής ανάλυσης διαφόρων μπλοκ πληροφοριών, την εισαγωγή νέων δεδομένων σε βάσεις δεδομένων υπολογιστών καθημερινά,
που λαμβάνονται σε διάφορα εργαστήρια σε όλο τον κόσμο και προσαρμόζουν τις νέες πληροφορίες σε αυτές που είχαν συσσωρευτεί νωρίτερα. Ταυτόχρονα, αναπτύχθηκαν συστήματα για την αποτελεσματική απομόνωση διαφόρων στοιχείων του γονιδιώματος και τον αυτόματο προσδιορισμό αλληλουχίας, δηλαδή τον προσδιορισμό των νουκλεοτιδικών αλληλουχιών του DNA. Σε αυτή τη βάση, σχεδιάστηκαν ισχυρά ρομπότ που επιταχύνουν σημαντικά την αλληλουχία και την καθιστούν λιγότερο δαπανηρή.
Η ανάπτυξη της γονιδιωματικής, με τη σειρά της, οδήγησε στην ανακάλυψη ενός τεράστιου αριθμού νέων γεγονότων. Η σημασία πολλών από αυτά μένει να εκτιμηθεί
μελλοντικός. Αλλά ακόμη και τώρα είναι προφανές ότι αυτές οι ανακαλύψεις θα οδηγήσουν σε μια επανεξέταση πολλών θεωρητικών θέσεων σχετικά με την εμφάνιση και την εξέλιξη διαφόρων μορφών ζωής στη Γη. Θα συμβάλουν στην καλύτερη κατανόηση των μοριακών μηχανισμών που διέπουν τη λειτουργία μεμονωμένων κυττάρων και τις αλληλεπιδράσεις τους. λεπτομερής αποκωδικοποίηση πολλών ακόμα άγνωστων βιοχημικών κύκλων.
ανάλυση της σύνδεσής τους με θεμελιώδεις φυσιολογικές διεργασίες.
Έτσι, υπάρχει μια μετάβαση από τη δομική γονιδιωματική σε
λειτουργικό, το οποίο με τη σειρά του δημιουργεί τις προϋποθέσεις για
έρευνα για τη μοριακή βάση της λειτουργίας των κυττάρων και του οργανισμού συνολικά.
Οι πληροφορίες που συγκεντρώνονται τώρα θα αποτελέσουν αντικείμενο ανάλυσης
τις επόμενες δεκαετίες. Αλλά σε κάθε επόμενο βήμα
κατεύθυνση της αποκρυπτογράφησης της δομής των γονιδιωμάτων διαφορετικών ειδών, δημιουργεί νέες τεχνολογίες που διευκολύνουν τη διαδικασία λήψης πληροφοριών. Ετσι,
η χρήση δεδομένων σχετικά με τη δομή και τη λειτουργία γονιδίων κατώτερων οργανωμένων ειδών έμβιων όντων μπορεί να επιταχύνει σημαντικά την αναζήτηση
αντικαθιστούν μάλλον απαιτητικές μοριακές μεθόδους για την αναζήτηση γονιδίων.
Η πιο σημαντική συνέπεια της αποκρυπτογράφησης της δομής του γονιδιώματος ενός συγκεκριμένου είδους είναι η ικανότητα αναγνώρισης όλων των γονιδίων του και
Κατά συνέπεια, ταυτοποίηση και προσδιορισμός της μοριακής φύσης των μεταγραμμένων μορίων RNA και όλων των πρωτεϊνών του. Κατ' αναλογία με το γονιδίωμα, γεννήθηκαν οι έννοιες του τρανσκριπτώματος, το οποίο ενώνει μια δεξαμενή μορίων RNA που σχηματίζονται ως αποτέλεσμα της μεταγραφής, και του ιπρωτεώματος, το οποίο περιλαμβάνει πολλές πρωτεΐνες που κωδικοποιούνται από γονίδια. Έτσι, η γονιδιωματική δημιουργεί τα θεμέλια για την εντατική ανάπτυξη νέων επιστημών – πρωτεομική και μεταγραφική. Πρωτεομική είναι η μελέτη της δομής και της λειτουργίας κάθε πρωτεΐνης. ανάλυση της πρωτεϊνικής σύνθεσης του κυττάρου. προσδιορισμός της μοριακής βάσης της λειτουργίας ενός μεμονωμένου κυττάρου, που είναι
το αποτέλεσμα της συντονισμένης εργασίας πολλών εκατοντάδων πρωτεϊνών, και
μελέτη του σχηματισμού ενός φαινοτυπικού χαρακτηριστικού ενός οργανισμού,
που προκύπτει από τη συντονισμένη εργασία δισεκατομμυρίων κυττάρων.
Πολύ σημαντικές βιολογικές διεργασίες συμβαίνουν επίσης σε επίπεδο RNA. Η ανάλυσή τους είναι αντικείμενο μεταγραφικής.
Οι μεγαλύτερες προσπάθειες επιστημόνων από πολλές χώρες του κόσμου που εργάζονται στον τομέα της γονιδιωματικής στόχευαν στην επίλυση του διεθνούς έργου «Human Genome». Σημαντική πρόοδος στον τομέα αυτό συνδέεται με την υλοποίηση της ιδέας,
που προτείνει ο J. S. Venter, αναζητήστε και αναλύστε
εκφρασμένες αλληλουχίες DNA, οι οποίες μπορούν αργότερα να χρησιμοποιηθούν ως ένα είδος «ετικέτες» ή δείκτες ορισμένων περιοχών του γονιδιώματος. Μια άλλη ανεξάρτητη και όχι λιγότερο γόνιμη προσέγγιση χρησιμοποιήθηκε στο έργο της ομάδας με επικεφαλής τον Fr.
Κόλινς. Βασίζεται στην πρωτογενή ταυτοποίηση γονιδίων για κληρονομικές ανθρώπινες ασθένειες.
Η αποκωδικοποίηση της δομής του ανθρώπινου γονιδιώματος οδήγησε σε μια συγκλονιστική ανακάλυψη. Αποδείχθηκε ότι το ανθρώπινο γονιδίωμα περιέχει μόνο 32.000 γονίδια, που είναι αρκετές φορές λιγότερο από τον αριθμό των πρωτεϊνών. Ταυτόχρονα, υπάρχουν μόνο 24.000 γονίδια που κωδικοποιούν πρωτεΐνες· τα προϊόντα των υπολοίπων γονιδίων είναι μόρια RNA.
Το ποσοστό ομοιότητας σε αλληλουχίες νουκλεοτιδίων DNA μεταξύ διαφορετικών ατόμων, εθνοτικών ομάδων και φυλών είναι 99,9%.
Αυτή η ομοιότητα είναι που μας κάνει ανθρώπους – Homo sapiens! Όλη η μεταβλητότητά μας σε επίπεδο νουκλεοτιδίων ταιριάζει σε ένα πολύ μέτριο ποσοστό - 0,1%.
Έτσι, η γενετική δεν αφήνει περιθώρια για ιδέες εθνικής ή φυλετικής ανωτερότητας.
Αλλά ας κοιτάξουμε ο ένας τον άλλον - είμαστε όλοι διαφορετικοί. Οι εθνικές, και πολύ περισσότερο, οι φυλετικές διαφορές είναι ακόμη πιο αισθητές. Ποιος λοιπόν αριθμός μεταλλάξεων καθορίζει την ανθρώπινη μεταβλητότητα, όχι σε ποσοστό, αλλά σε απόλυτες τιμές; Για να λάβετε αυτήν την εκτίμηση, πρέπει να θυμάστε ποιο είναι το μέγεθος του γονιδιώματος. Το μήκος ενός ανθρώπινου μορίου DNA είναι
3,2x109 ζεύγη βάσεων. Το 0,1% αυτού είναι 3,2 εκατομμύρια νουκλεοτίδια. Αλλά να θυμάστε ότι το κωδικοποιητικό μέρος του γονιδιώματος καταλαμβάνει λιγότερο από το 3% του συνολικού μήκους του μορίου DNA και οι μεταλλάξεις εκτός αυτής της περιοχής, τις περισσότερες φορές, δεν έχουν καμία επίδραση στη φαινοτυπική μεταβλητότητα. Έτσι, για να λάβουμε μια ολοκληρωμένη εκτίμηση του αριθμού των μεταλλάξεων που επηρεάζουν τον φαινότυπο, πρέπει να πάρουμε το 3% των 3,2 εκατομμυρίων νουκλεοτιδίων, που θα μας δώσει έναν αριθμό της τάξης των 100.000. Δηλαδή, περίπου 100 χιλιάδες μεταλλάξεις αποτελούν το φαινοτυπικό μας μεταβλητότητα. Εάν συγκρίνουμε αυτόν τον αριθμό με τον συνολικό αριθμό γονιδίων, αποδεικνύεται ότι κατά μέσο όρο υπάρχουν 3-4 μεταλλάξεις ανά γονίδιο.
Ποιες είναι αυτές οι μεταλλάξεις; Η συντριπτική τους πλειοψηφία (τουλάχιστον 70%)
καθορίζει την ατομική μας μη παθολογική μεταβλητότητα, αυτό που μας διακρίνει, αλλά δεν μας κάνει χειρότερους μεταξύ μας. Αυτό περιλαμβάνει χαρακτηριστικά όπως χρώμα ματιών, μαλλιά, δέρμα, σωματότυπο, ύψος, βάρος,
ένας τύπος συμπεριφοράς που επίσης καθορίζεται σε μεγάλο βαθμό γενετικά, και πολλά άλλα. Περίπου το 5% των μεταλλάξεων σχετίζονται με μονογονιδιακά νοσήματα. Περίπου το ένα τέταρτο των υπόλοιπων μεταλλάξεων ανήκουν στην κατηγορία των λειτουργικών πολυμορφισμών. Συμμετέχουν στο σχηματισμό κληρονομικής προδιάθεσης για εκτεταμένη πολυπαραγοντική παθολογία. Φυσικά, αυτές οι εκτιμήσεις είναι πολύ σκληρές,
αλλά καθιστούν δυνατή την κρίση της δομής της ανθρώπινης κληρονομικής μεταβλητότητας.
Κεφάλαιο 1.16. Μοριακή γενετική βάση της εξέλιξης
Η επανάσταση στον τομέα της μοριακής βιολογίας που συνέβη στις αρχές της χιλιετίας, με αποκορύφωμα την αποκρυπτογράφηση της δομής των γονιδιωμάτων πολλών εκατοντάδων ειδών μικροοργανισμών, καθώς και ορισμένων ειδών πρωτοζώων,
μαγιά, φυτά, ζώα και άνθρωποι, ανέτρεψαν πολλές παραδοσιακές ιδέες της κλασικής γενετικής και έφεραν τη δυνατότητα μελέτης των μοριακών μηχανισμών της εξέλιξης και της ειδογένεσης πολύ κοντά. Μια νέα επιστήμη γεννήθηκε - η συγκριτική γονιδιωματική,
καθιστώντας δυνατή την καταγραφή της εμφάνισης σε διάφορες φυλογενετικές γραμμές εξελικτικά σημαντικών γεγονότων που συμβαίνουν σε επίπεδο μεμονωμένων μορίων. Αποδείχθηκε ότι στη γενική περίπτωση, η εξελικτική πρόοδος συνδέεται όχι μόνο, και όχι τόσο με την αύξηση του αριθμού, της έκτασης και ακόμη και της πολυπλοκότητας της δομικής οργάνωσης των γονιδίων, αλλά σε πολύ μεγαλύτερο βαθμό με μια αλλαγή στον κανονισμό του έργου τους, το οποίο καθορίζει τον συντονισμό και την εξειδίκευση των ιστών της έκφρασης δεκάδων χιλιάδων γονιδίων. Αυτό, τελικά, οδήγησε στην εμφάνιση σε ανώτερους οργανισμούς πιο πολύπλοκων, ιδιαίτερα ειδικών, πολυλειτουργικών συμπλεγμάτων αλληλεπιδρώντων πρωτεϊνών ικανών να εκτελούν θεμελιωδώς νέες εργασίες.
Ας εξετάσουμε τη φύση των αλλαγών που συμβαίνουν στη διαδικασία της εξέλιξης σε τρία επίπεδα πληροφοριών: DNA - RNA - πρωτεΐνη ή γονιδίωμα - μεταγραφή - πρωτεόμιο. Γενικά, μπορούμε να πούμε ότι όσο αυξάνεται η πολυπλοκότητα της οργάνωσης της ζωής, τόσο το μέγεθος του γονιδιώματος αυξάνεται. Έτσι, το μέγεθος του DNA των προκαρυωτών δεν υπερβαίνει τα 8x106 bp, γίνεται διπλάσιο σε ζυμομύκητες και πρωτόζωα, 10-15 φορές μεγαλύτερο στα έντομα και στα θηλαστικά η αύξηση φτάνει τις 3 τάξεις μεγέθους, δηλαδή χίλιες φορές (103 ).
Ωστόσο, αυτή η εξάρτηση δεν είναι γραμμική. Έτσι, στα θηλαστικά δεν παρατηρούμε πλέον σημαντική αύξηση στο μέγεθος του γονιδιώματος. Επιπλέον, δεν είναι πάντα δυνατό να παρατηρηθεί η σχέση μεταξύ του μεγέθους του γονιδιώματος και της πολυπλοκότητας της οργάνωσης της ζωής. Έτσι, σε ορισμένα φυτά το μέγεθος του γονιδιώματος είναι μια τάξη μεγέθους ή ακόμη και δύο τάξεις μεγέθους μεγαλύτερο από αυτό των ανθρώπων. Ας θυμηθούμε ότι η αύξηση του μεγέθους του γονιδιώματος των ευκαρυωτών σε σύγκριση με τα προκαρυωτικά συμβαίνει κυρίως λόγω της εμφάνισης μη κωδικοποιητικών αλληλουχιών, δηλαδή προαιρετικών στοιχείων. Έχουμε ήδη πει ότι στο ανθρώπινο γονιδίωμα τα εξόνια συνολικά δεν υπερβαίνουν το 1-3%. Αυτό σημαίνει ότι ο αριθμός των γονιδίων σε ανώτερους οργανισμούς μπορεί να είναι μόνο αρκετές φορές μεγαλύτερος από ό,τι στους μικροοργανισμούς.
Η αυξανόμενη πολυπλοκότητα της ευκαρυωτικής οργάνωσης εξηγείται εν μέρει από την εμφάνιση ενός πρόσθετου ρυθμιστικού συστήματος που είναι απαραίτητο για
διασφαλίζοντας την εξειδίκευση των ιστών της γονιδιακής έκφρασης. Μία από τις συνέπειες της ασυνεχούς οργάνωσης των γονιδίων που εμφανίστηκαν στους ευκαρυώτες ήταν η ευρεία εμφάνιση εναλλακτικού ματίσματος και εναλλακτικής μεταγραφής. Αυτό οδήγησε στην εμφάνιση μιας νέας ιδιότητας σε έναν τεράστιο αριθμό γονιδίων - την ικανότητα κωδικοποίησης πολλαπλών λειτουργικά διαφορετικών ισομορφών πρωτεΐνης. Έτσι, η συνολική ποσότητα πρωτεϊνών
δηλαδή το μέγεθος του πρωτεώματος· οι υψηλότερες μπορεί να έχουν πολλαπλάσιο αριθμό γονιδίων.
Στους προκαρυώτες, η ενδοειδική μεταβλητότητα στον αριθμό των γονιδίων είναι επιτρεπτή και
παρόμοιες διαφορές μεταξύ διαφορετικών στελεχών πολλών μικροοργανισμών, σε
συμπεριλαμβανομένων των παθογόνων, μπορεί να ανέλθει σε δεκάδες τοις εκατό. Επιπλέον, η πολυπλοκότητα της οργάνωσης διαφόρων τύπων μικροοργανισμών συσχετίζεται άμεσα με τον αριθμό και το μήκος των κωδικοποιητικών αλληλουχιών.
Έτσι, η φαινοτυπική ενδοειδική και ενδοειδική μεταβλητότητα είναι σε αυστηρή συσχέτιση με πολύ παρόμοια μεγέθη μεταγραφών και πρωτεωμάτων. Στους ευκαρυώτες, ο αριθμός των γονιδίων είναι ένα αυστηρά καθορισμένο χαρακτηριστικό του είδους και η αύξηση της εξελικτικής πολυπλοκότητας βασίζεται σε μια άλλη αρχή - τη διαφορική πολυεπίπεδη χρήση διαφόρων συστατικών ενός περιορισμένου και αρκετά σταθερού πρωτεώματος.
Η αλληλούχιση των γονιδιωμάτων των νηματωδών και της Drosophila έδειξε ότι τα μεγέθη των πρωτεωμάτων σε αυτά τα πολύ διαφορετικά είδη είναι πολύ παρόμοια και μόνο δύο φορές μεγαλύτερα από αυτά της ζύμης και ορισμένων τύπων βακτηρίων. Αυτό το μοτίβο - μια σημαντική αύξηση στην πολυπλοκότητα της οργάνωσης των διαφόρων μορφών ζωής με παράλληλη διατήρηση ή μια σχετικά μικρή αύξηση στο μέγεθος του πρωτεώματος - είναι χαρακτηριστικό όλων των επακόλουθων εξελίξεων μέχρι τον άνθρωπο. Ετσι,
Τα πρωτεώματα των ανθρώπων και των ποντικών ουσιαστικά δεν διαφέρουν μεταξύ τους και είναι λιγότερο από 2 φορές μεγαλύτερα σε μέγεθος από τα πρωτεώματα του μικροσκοπικού νηματώδους στρογγυλού σκουληκιού ή της μύγας των φρούτων Drosophila. Επιπλέον, η ταυτότητα των νουκλεοτιδικών αλληλουχιών του ανθρώπινου DNA και
μεγάλοι πίθηκοι είναι 98,5%, και στις περιοχές κωδικοποίησης φτάνει το 99%. Αυτά τα στοιχεία διαφέρουν ελάχιστα από την τιμή του 99,9%,
τον προσδιορισμό της ενδοειδικής ομοιότητας στις αλληλουχίες νουκλεοτιδίων του DNA μεταξύ διαφορετικών ατόμων, λαών και φυλών που κατοικούν στον πλανήτη μας. Ποιες αλλαγές λοιπόν, που δεν αποτελούν περισσότερο από το 1,5% ολόκληρου του γονιδιώματος, είναι βασικές για το σχηματισμό ενός ατόμου; Η απάντηση σε αυτό το ερώτημα, προφανώς, θα πρέπει να αναζητηθεί όχι μόνο στο γονιδιωματικό και πρωτεομικό επίπεδο.
Πράγματι, μαζί με τη σχετική σταθερότητα του πρωτεώματος, στο
Στη διαδικασία της εξέλιξης, παρατηρείται μια απότομη αύξηση στο μέγεθος και την πολυπλοκότητα της οργάνωσης του ευκαρυωτικού μεταγραφώματος λόγω της εμφάνισης στο γονιδίωμα ενός τεράστιου αριθμού μεταγραμμένων και μη κωδικοποιημένων DNA, καθώς και σημαντική επέκταση του κατηγορία γονιδίων που κωδικοποιούν το RNA. RNA που δεν κωδικοποιούν πρωτεΐνες, η κύρια πηγή των οποίων είναι τα ιντρόνια,
αποτελούν τη συντριπτική πλειοψηφία του μεταγραφήματος των ανώτερων οργανισμών,
φτάνοντας το 97-98% όλων των μεταγραφικών μονάδων. Οι λειτουργίες αυτών των μορίων αναλύονται επί του παρόντος εντατικά.
Έτσι, βασικές εξελικτικές αλλαγές συμβαίνουν στο πλαίσιο μιας αύξησης του μεγέθους του γονιδιώματος, ενός αρκετά σταθερού πρωτεώματος και μιας απότομης αύξησης στο μέγεθος του μεταγραφώματος - Εικ. 31.
Εικόνα 31. Εξελικτικές αλλαγές που συμβαίνουν σε τρία
επίπεδα πληροφόρησης Ταυτόχρονα, η μετάβαση από απλές μορφές ζωής σε πιο σύνθετες είναι εμφανής
συσχετίζεται με την εμφάνιση και την ευρεία κατανομή στο γονιδίωμα δύο θεμελιωδών και σε κάποιο βαθμό αλληλένδετων εξελικτικών αποκτήσεων: του μη κωδικοποιητικού DNA και των επαναλαμβανόμενων στοιχείων. Άμεση συνέπεια αυτών των αλλαγών που συμβαίνουν σε γονιδιωματικό επίπεδο είναι η εμφάνιση στη διαδικασία της εξέλιξης ενός τεράστιου αριθμού RNA που δεν κωδικοποιούν πρωτεΐνες.
Ποια είναι η δομική βάση αυτών των εξελικτικών μετασχηματισμών;
Όλες οι κύριες εξελικτικές μεταβάσεις: από προκαρυώτες στους ευκαρυώτες, από πρωτόζωα στα μεταζώα, από τα πρώτα ζώα στα διστεροειδή και από τα πρωτόγονα χορδοειδή στα σπονδυλωτά, συνοδεύτηκαν από μια απότομη αύξηση της πολυπλοκότητας του γονιδιώματος. Προφανώς, τέτοια άλματα στην εξέλιξη είναι το αποτέλεσμα σπάνιων περιπτώσεων επιτυχούς σύντηξης ολόκληρων γονιδιωμάτων διαφορετικών ειδών που ανήκουν σε συστηματικές τάξεις που έχουν αποκλίνει σε σημαντική απόσταση το ένα από το άλλο. Έτσι, η συμβίωση των Αρχαίων και των Βακτηρίων σηματοδότησε την αρχή της μετάβασης από τους προκαρυώτες στους ευκαρυώτες. Είναι προφανές ότι ως αποτέλεσμα της ενδοσυμβίωσης εμφανίστηκαν και μιτοχόνδρια, χλωροπλάστες και κάποια άλλα κυτταρικά οργανίδια. Μια θεμελιώδης ιδιότητα των ανώτερων ευκαρυωτών, η διπλοειδία, προέκυψε ως αποτέλεσμα του καλά ρυθμισμένου γονιδιωματικού διπλασιασμού που συνέβη πριν από περίπου 500 εκατομμύρια χρόνια.
Οι διπλασιασμοί του γονιδιώματος μέσα σε ένα είδος συνέβησαν αρκετά συχνά, και
Παραδείγματα αυτού είναι πολλές περιπτώσεις πολυπλοειδίας σε φυτά,
μανιτάρια και μερικές φορές ακόμη και σε ζώα. Ωστόσο, πιθανοί μηχανισμοί
που οδηγούν στην εμφάνιση θεμελιωδώς νέων μορφών ζωής στη διαδικασία της εξέλιξης δεν είναι η αυτοπολυπλοειδία, αλλά ο υβριδισμός και η οριζόντια μεταφορά ή σύντηξη γονιδιωμάτων. Αξίζει να σημειωθεί ότι οι πιο σημαντικοί εξελικτικοί μετασχηματισμοί, που συνοδεύονται από τη σύντηξη ολόκληρων γονιδιωμάτων, συμβαίνουν κάτω από εξαιρετικές συνθήκες, κατά τη διάρκεια περιόδων μεγάλων γεωλογικών μεταβάσεων, όπως αλλαγές στη συγκέντρωση οξυγόνου στην ατμόσφαιρα, παγετώνες της Γης ή της Κάμβριας Εκρηξη.
Σε σχετικά ήρεμες γεωλογικές συνθήκες, οι διπλασιασμοί μεμονωμένων γονιδίων ή χρωμοσωμικών τμημάτων με την επακόλουθη απόκλιση αποδεικνύονται πιο σημαντικές για την εξέλιξη. Μια σύγκριση των αλληλουχιών νουκλεοτιδίων των γονιδιωμάτων με αλληλουχία δείχνει ότι η συχνότητα των διπλασιασμών γονιδίων είναι αρκετά υψηλή και, κατά μέσο όρο, είναι 0,01 ανά γονίδιο ανά εκατομμύριο χρόνια. Η συντριπτική πλειονότητά τους δεν εκδηλώνεται κατά τα επόμενα πολλά εκατομμύρια χρόνια, και μόνο σε σπάνιες περιπτώσεις
Σε περιπτώσεις, τα διπλά γονίδια μπορούν να αποκτήσουν νέες προσαρμοστικές λειτουργίες. Ωστόσο, μια μεγάλη κατηγορία «σιωπηλών» αντιγραφών γονιδίων χρησιμεύει ως ένα είδος αποθεματικού ταμείου για τη γέννηση νέων γονιδίων και το σχηματισμό νέων ειδών. Το ανθρώπινο γονιδίωμα περιέχει από 10 έως 20 χιλιάδες αντίγραφα επεξεργασμένων γονιδίων που προέκυψαν μέσω της επανατοποθέτησης του mRNA.
Τα περισσότερα από αυτά ανήκουν στην κατηγορία των ψευδογονιδίων, δηλαδή δεν εκφράζονται ούτε λόγω παρουσίας μεταλλάξεων ούτε λόγω εισαγωγών σε μεταγραφικά ανενεργές περιοχές του γονιδιώματος. Ωστόσο, ορισμένα από αυτά τα γονίδια είναι ενεργά και η φύση της έκφρασης και ακόμη και οι λειτουργίες τους μπορεί να είναι διαφορετικές,
από εκείνα των ιδρυτικών γονιδίων.
Παίζουν ιδιαίτερο ρόλο στην εξέλιξη των πρωτευόντων και των ανθρώπων. τμηματικές αντιγραφές, που ανήκουν στην κατηγορία των χαμηλών επαναλήψεων αντιγραφής (LCR) και
εμφανίστηκε πριν από λιγότερο από 35 εκατομμύρια χρόνια. Αυτές οι αλληλουχίες είναι πανομοιότυπα μπλοκ DNA, που ποικίλλουν σε μέγεθος από μία έως αρκετές εκατοντάδες κιλοβάσεις. Τις περισσότερες φορές, οι τμηματικοί διπλασιασμοί εντοπίζονται στις περικεντρομερείς ή τελομερείς περιοχές διαφόρων χρωμοσωμάτων και συνολικά καταλαμβάνουν περίπου το 5% του ανθρώπινου γονιδιώματος.
Δεν βρέθηκαν τμηματικοί διπλασιασμοί σε άλλα γονιδιώματα με αλληλουχία.
Η ελάχιστη ενότητα τμηματικού διπλασιασμού, που ονομάζεται διπλότυπο, περιέχει θραύσματα άσχετων μη επεξεργασμένων γονιδίων και
Αυτό το διακρίνει από άλλους γνωστούς τύπους επαναλαμβανόμενων αλληλουχιών. Κάτω από ορισμένες συνθήκες, τα διπλάσια μπορούν να χρησιμεύσουν ως πηγές για τη δημιουργία νέων χιμαιρικών μεταγραφόμενων γονιδίων ή οικογενειών γονιδίων από διάφορους συνδυασμούς κωδικοποιητικών εξονίων που υπάρχουν σε αυτά. Υπολογίζεται ότι μεταξύ 150 και 350 γονιδίων μπορούν να διακρίνουν το γονιδίωμα του χιμπατζή και του ανθρώπου.
Χωρίς να μειώνουμε τη σημασία της εμφάνισης νέων και της εξαφάνισης των παλαιών κωδικοποιητικών ακολουθιών για την ειδογένεση, θα πρέπει να τονίσουμε την πραγματική πιθανότητα ύπαρξης άλλων μηχανισμών,
παίζοντας καθοριστικό ρόλο στην εξέλιξη των ευκαρυωτών.
Ένας από τους κινητήριους μηχανισμούς της εξέλιξης είναι τα κινητά στοιχεία, που βρίσκονται σε όλα τα είδη που μελετήθηκαν από αυτή την άποψη.
Οι γονιδιωματικές αλλαγές που συνοδεύουν τη διαδικασία της ειδογένεσης μπορεί να περιλαμβάνουν εκτεταμένες αναδιοργανώσεις καρυότυπου, τοπικές χρωμοσωμικές ανακατατάξεις, διπλασιασμούς οικογενειών γονιδίων, τροποποιήσεις μεμονωμένων γονιδίων,
συνοδεύεται από τη γέννηση ή την απώλειά τους, καθώς και από διαφορές στη γονιδιακή έκφραση, που ρυθμίζονται τόσο σε επίπεδο μεταγραφής όσο και σε επίπεδα ματίσματος ή μετάφρασης. Τα κινητά στοιχεία σχετίζονται άμεσα με όλες αυτές τις διαδικασίες.
Σε ορισμένες περιπτώσεις, τα ίδια τα μετατιθέμενα στοιχεία φέρουν αλληλουχίες που κωδικοποιούν ένζυμα των οποίων η παρουσία είναι απαραίτητη για τη μεταφορά DNA ή την αναδρομή RNA.
Παρόμοιες αλληλουχίες υπάρχουν στο γονιδίωμα των ρετροϊών, LTR-
στοιχεία και τρανσποζόνια. Τα ρετροτρανσποζόνια περιλαμβάνουν επίσης την πιο πολυάριθμη κατηγορία μετατιθέμενων στοιχείων - τις επαναλήψεις Alu. Για πρώτη φορά Alu-
επαναλήψεις εμφανίζονται στα πρωτεύοντα περίπου 50-60 εκατομμύρια χρόνια πριν από ένα μικρό γονίδιο που κωδικοποιεί το RNA. Στη διαδικασία της περαιτέρω εξέλιξης, εμφανίζεται απόκλιση και ισχυρή ενίσχυση αυτής της οικογένειας. Η μετάβαση από τα πρωτεύοντα στον άνθρωπο συνοδεύεται από εκρηκτική αύξηση του αριθμού
Το Alu επαναλαμβάνει, ο αριθμός των αντιγράφων των οποίων, σύμφωνα με ορισμένες εκτιμήσεις, φτάνει
1,1 εκατ. Οι επαναλήψεις του Alu καταλαμβάνουν περίπου το 10% του ανθρώπινου γονιδιώματος, αλλά η κατανομή τους είναι άνιση, καθώς συνδέονται κυρίως με γονίδια. Αυτά τα στοιχεία σπάνια υπάρχουν στα κωδικοποιητικά εξόνια και απαντώνται αρκετά συχνά σε ιντρόνια και μη κωδικοποιητικές περιοχές του mRNA, επηρεάζοντας τη σταθερότητα αυτών των μορίων ή/και την αποτελεσματικότητα της μετάφρασης. Η παρουσία αλληλουχιών Alu στις ιντρονικές περιοχές των γονιδίων μπορεί να συνοδεύεται από μια αλλαγή στη φύση της επεξεργασίας του preRNA, καθώς αυτές οι αλληλουχίες περιέχουν περιοχές ομόλογες με τις θέσεις ματίσματος δότη και δέκτη. Όταν τα στοιχεία Alu εισάγονται στις ρυθμιστικές περιοχές ενός γονιδίου, η μεταγραφή μπορεί να διαταραχθεί, με αποτέλεσμα
Μια ομάδα ερευνητών από τα Πανεπιστήμια του Σικάγο, του Ουισκόνσιν και της Νεμπράσκα-Λίνκολν διέψευσε μια από τις κλασικές υποθέσεις της εξελικτικής προσαρμογής χρησιμοποιώντας το παράδειγμα της φρουτόμυγας Drosophila vulgaris (Drosophila). Drosophila melanogaster). Τον 20ο αιώνα, αυτές οι μύγες έγιναν ένα κύριο πρότυπο οργανισμό για την αναπτυξιακή βιολογία και έκτοτε χρησιμοποιούνται συχνά σε γενετικά πειράματα.
Οι επιστήμονες συνέθεσαν μια αρχαία εκδοχή ενός από τα γονίδια του είδους και δημιούργησαν διαγονιδιακά έντομα (δηλαδή με γονίδια που δεν μπορούν να αποκτηθούν μέσω φυσικής διασταύρωσης). Πειράματα μαζί τους έδειξαν ότι η προσαρμογή των μυγών στις μεταβαλλόμενες περιβαλλοντικές συνθήκες κατά την ανάπτυξη του είδους δεν συνέβη όπως πιστεύαμε προηγουμένως. (Ένα συγκεκριμένο παράδειγμα με επεξήγηση θα δοθεί παρακάτω.)
"Ένας από τους κύριους στόχους της σύγχρονης εξελικτικής βιολογίας είναι να προσδιορίσει ποια γονίδια επέτρεψαν τα είδη. Αλλά αυτό είναι δύσκολο να γίνει άμεσα γιατί δεν μπορούμε να ελέγξουμε την επιρροή των αρχαίων γονιδίων στη βιολογία των ζώων", λέει ο Mo Siddiq του Πανεπιστημίου του Σικάγο. «Συνειδητοποιήσαμε ότι μπορούσαμε να λύσουμε αυτό το πρόβλημα χρησιμοποιώντας δύο τεχνικές που αναπτύχθηκαν πρόσφατα - τη στατιστική ανακατασκευή αρχαίων αλληλουχιών νουκλεοτιδίων και τη δημιουργία διαγονιδιακών οργανισμών».
Με απλά λόγια, οι βιολόγοι αποφάσισαν πρώτα να «γυρίσουν πίσω το ρολόι» για να καταλάβουν πώς ήταν τα γονίδια της μύγας φρούτων πριν, στη συνέχεια να συνθέσουν ένα από αυτά και να το εισαγάγουν στο γονιδίωμα του εντόμου για να δουν πώς θα άλλαζε η ζωή του.
Όταν οι επιστήμονες μελετούν την προσαρμογή σε μοριακό επίπεδο, η ανάλυση των αλληλουχιών νουκλεοτιδίων στο DNA τους βοηθά να εντοπίσουν τις λεγόμενες υπογραφές επιλογής, οι οποίες υποδεικνύουν γρήγορες αλλαγές σε ένα γονίδιο στο παρελθόν. Ωστόσο, τέτοια στοιχεία γραμμένα στο γονιδίωμα μπορούν να θεωρηθούν μόνο έμμεσα, επειδή μπορεί να υπάρχουν πολλοί λόγοι για την εξέλιξη των γονιδίων και δεν σχετίζονται απαραίτητα με τις μεταβαλλόμενες περιβαλλοντικές συνθήκες στις οποίες έχει προσαρμοστεί ο οργανισμός. Συνεπώς, η πρώτη μέθοδος που αναφέρθηκε παραπάνω από τον Siddique δεν είναι επαρκής.
Σε μια νέα μελέτη, οι επιστήμονες προσπάθησαν να αξιολογήσουν άμεσα την επίδραση της εξέλιξης των γονιδίων στην προσαρμογή, προσθέτοντας μια δεύτερη μέθοδο στην πρώτη. Ο επικεφαλής της εργασίας, Joe Thornton, σε προηγούμενες μελέτες του είχε ήδη χρησιμοποιήσει τη μέθοδο της στατιστικής ανακατασκευής αλληλουχιών νουκλεοτιδίων, βασιζόμενος σε εκτεταμένες βάσεις δεδομένων για τη δομή των γονιδιωμάτων των σύγχρονων οργανισμών, τα συνέθεσε και ανέλυσε τις μοριακές ιδιότητες των γονιδίων που προέκυψαν. στο εργαστήριο.
Πρότεινε ότι η γενετική μηχανική και η ανακατασκευή αρχαίων γονιδίων μαζί θα μπορούσαν να δείξουν πώς οι αλλαγές στο γονιδίωμα θα μπορούσαν να επηρεάσουν τον οργανισμό ως σύνολο.
"Αυτή η τεχνική δημιουργίας μιας αρχαίας εκδοχής ζώων μπορεί να εφαρμοστεί στη μελέτη διαφορετικών πτυχών της εξέλιξης", σημειώνει ο Thornton. "Για το πρώτο πείραμα, επιλέξαμε ένα κλασικό παράδειγμα προσαρμογής - μια μύγα φρούτων, η οποία κατά τη διαδικασία της εξέλιξης απέκτησε την ικανότητα που περιέχεται στα φρούτα που σαπίζουν Βρήκαμε ότι ευρέως διαδεδομένη «Η υπόθεση για τους μηχανισμούς εξέλιξης των φρουτόμυγων είναι απλά λανθασμένη».
Ας εξηγήσουμε τι υπάρχει στην άγρια φύση D. melanogasterτρέφεται με προϊόντα ζύμωσης φρούτων (μικρά έντομα εμφανίζονται συχνά και πολλαπλασιάζονται γρήγορα εκεί που βρίσκονται τα χτυπημένα φρούτα). Είναι σε θέση να ανεχθούν υψηλότερες συγκεντρώσεις αλκοόλ από τους στενότερους συγγενείς τους, που τρέφονται με άλλα τρόφιμα.
Πριν από είκοσι πέντε χρόνια, οι βιολόγοι Martin Kreitman και John McDonald του Πανεπιστημίου του Σικάγο ανέπτυξαν τη στατιστική τους μέθοδο για τον εντοπισμό των υπογραφών επιλογής, η οποία παραμένει μια από τις πιο ευρέως χρησιμοποιούμενες μεθόδους σήμερα. Έδειξαν την εγκυρότητά του χρησιμοποιώντας το παράδειγμα του γονιδίου της αφυδρογονάσης της αλκοόλης (ADH). Αυτό το γονίδιο κωδικοποιεί ένα ένζυμο που διασπά την αιθανόλη στα ηπατικά κύτταρα.
Οι Kreitman και McDonald βρήκαν ίχνη επιλογής στην αλληλουχία νουκλεοτιδίων της ADH, και αφού γνώριζαν ότι το σώμα της μύγας διασπά τις αλκοόλες πιο γρήγορα από τους συγγενείς της, οι επιστήμονες πρότειναν ότι ήταν το ένζυμο που κωδικοποιήθηκε από την ADH που βοήθησε τη μύγα να προσαρμοστεί στο υψηλές συγκεντρώσεις αλκοόλ. Ως αποτέλεσμα, αυτή η εργασία έγινε η πρώτη περίπτωση που αναγνωρίστηκε από την επιστημονική κοινότητα στην οποία ένα συγκεκριμένο γονίδιο επηρέασε την προσαρμοστική εξέλιξη ενός είδους.
Τώρα ερευνητές από τις Ηνωμένες Πολιτείες αποφάσισαν να δοκιμάσουν την υπόθεση χρησιμοποιώντας άλλες τεχνολογίες. Μελέτησαν παραλλαγές στις νουκλεοτιδικές αλληλουχίες του γονιδίου πριν και μετά την απόκτηση της ανοχής στην αιθανόλη από τη μύγα των φρούτων (περίπου 2-4 εκατομμύρια χρόνια πριν). Στη συνέχεια οι επιστήμονες συνέθεσαν αυτά τα γονίδια, ξεκίνησαν την έκφρασή τους και εξέτασαν την ικανότητα των πρωτεϊνών που προέκυψαν να διασπούν τις αλκοόλες. Ως αποτέλεσμα, αποδείχθηκε ότι οι αλλαγές που υπέστη το γονιδίωμα της μύγας των φρούτων κατά την εξέλιξη δεν είχαν ιδιαίτερο αντίκτυπο στη λειτουργία του ενζύμου.
Στη συνέχεια, οι επιστήμονες εισήγαγαν μια «αρχαία» μορφή του γονιδίου ADH στο γονιδίωμα των σύγχρονων μυγών φρούτων και εξέτρεψαν χιλιάδες τροποποιημένα έντομα για να δοκιμάσουν πόσο γρήγορα θα διασπούσαν την αιθανόλη και πόσο θα μπορούσαν να επιβιώσουν εάν τρέφονταν με σάπια φρούτα με υψηλή περιεκτικότητα σε αλκοόλ. .
Τα πειράματα έδειξαν ότι οι μύγες των φρούτων με μια παλαιότερη μορφή του γονιδίου ADH επεξεργάζονταν την αιθανόλη όχι χειρότερα από τα έντομα με την «τελευταία έκδοση» του γονιδίου. Επιπλέον, μεγάλωσαν και αναπαράγονταν με τον ίδιο τρόπο, τρώγοντας τρόφιμα με υψηλή περιεκτικότητα σε αλκοόλ.
Έτσι, η κλασική υπόθεση δεν επιβεβαιώθηκε, ακριβέστερα, η μορφή D. melanogasterόντως προσαρμόστηκε σε τρόφιμα πλούσια σε αλκοόλες κατά τη διαδικασία της εξέλιξης, αλλά οι αλλαγές στο ένζυμο αφυδρογονάση της αλκοόλης δεν συνδέονται με αυτό, συμπεραίνουν οι επιστήμονες.
Όπως εξηγεί ο Thornton, η υπόθεση του γονιδίου ADH έγινε αποδεκτή εκείνη την εποχή, επειδή η οικολογία, η φυσιολογία και τα στατιστικά στοιχεία επιλογής έδειχναν προς την ίδια κατεύθυνση.
"Αλλά τα περιστασιακά στοιχεία δεν σημαίνουν ότι μια υπόθεση είναι απαραίτητα σωστή. Γι' αυτό, τώρα που η τεχνολογία μας έδωσε αυτή την ευκαιρία, θέλαμε να δοκιμάσουμε άμεσα αυτήν την υπόθεση", προσθέτει.
Μια ομάδα αμερικανών ερευνητών ελπίζει ότι η νέα τεχνική για την απόκτηση οργανισμών με πιο αρχαίες εκδόσεις γονιδίων θα γίνει το λεγόμενο χρυσό πρότυπο σε αυτόν τον τομέα και στο μέλλον θα βοηθήσει στον προσδιορισμό ακριβώς ποιες γενετικές αλλαγές επηρέασαν τα εξελικτικά χαρακτηριστικά των οργανισμών.
Το έργο των Αμερικανών γενετιστών περιγράφεται με περισσότερες λεπτομέρειες σε επιστημονικό άρθρο που δημοσιεύτηκε στο περιοδικό Nature Ecology & Evolution.
Ας θυμηθούμε ότι προηγουμένως μιλήσαμε για το πώς τα φύκια βαθέων υδάτων ανάγκασαν τους επιστήμονες να υποστούν εξελικτική προσαρμογή. Επιπλέον, οι δυνατότητες της εξαιρετικά γρήγορης εξέλιξης έχουν πρόσφατα αποδειχθεί από το .
Εκδοτικός οίκος «ΜΠΙΝΟΜ. Το Knowledge Laboratory κυκλοφορεί ένα βιβλίο με απομνημονεύματα του γενετιστή Craig Venter, Life Deciphered. Ο Craig Venter είναι γνωστός για το έργο του σχετικά με την ανάγνωση και την αποκρυπτογράφηση του ανθρώπινου γονιδιώματος. Το 1992 ίδρυσε το Ινστιτούτο Έρευνας Γονιδιώματος (TIGR). Το 2010, ο Venter δημιούργησε τον πρώτο τεχνητό οργανισμό στον κόσμο - το συνθετικό βακτήριο Mycoplasma εργαστήριο. Σας προσκαλούμε να διαβάσετε ένα από τα κεφάλαια του βιβλίου, στο οποίο ο Craig Venter μιλά για το έργο του 1999–2000 για την αλληλουχία του γονιδιώματος της μύγας Drosophila.
Εμπρός και μόνο μπροστά
Οι θεμελιώδεις πτυχές της κληρονομικότητας αποδείχθηκαν, προς έκπληξή μας, αρκετά απλές, και επομένως υπήρχε ελπίδα ότι ίσως η φύση δεν είναι τόσο άγνωστη και η ακατανόητη, που διακηρύσσεται επανειλημμένα από διάφορους ανθρώπους, είναι απλώς μια άλλη ψευδαίσθηση, ο καρπός της άγνοιάς μας . Αυτό μας κάνει αισιόδοξους, γιατί αν ο κόσμος ήταν τόσο περίπλοκος όσο ισχυρίζονται ορισμένοι φίλοι μας, η βιολογία δεν θα είχε καμία πιθανότητα να γίνει ακριβής επιστήμη.
Τόμας Χαντ Μόργκαν. Φυσική βάση της κληρονομικότητας
Πολλοί άνθρωποι με έχουν ρωτήσει γιατί, από όλα τα ζωντανά πλάσματα στον πλανήτη μας, επέλεξα τη μύγα. άλλοι αναρωτήθηκαν γιατί δεν προχώρησα αμέσως στην αποκρυπτογράφηση του ανθρώπινου γονιδιώματος. Το θέμα είναι ότι χρειαζόμασταν μια βάση για μελλοντικά πειράματα, θέλαμε να είμαστε σίγουροι για την ορθότητα της μεθόδου μας προτού δαπανήσουμε σχεδόν 100 εκατομμύρια δολάρια για τον προσδιορισμό της αλληλουχίας του ανθρώπινου γονιδιώματος.
Η μικρή μύγα έπαιξε τεράστιο ρόλο στην ανάπτυξη της βιολογίας, ιδιαίτερα της γενετικής. Το γένος Drosophila περιλαμβάνει διάφορες μύγες - ξύδι, κρασί, μήλο, σταφύλι και φρούτα - συνολικά περίπου 26 εκατοντάδες είδη. Αλλά πείτε τη λέξη "drosophila" και οποιοσδήποτε επιστήμονας θα σκεφτεί αμέσως ένα συγκεκριμένο είδος - το Drosophilamelanogaster. Επειδή αναπαράγεται γρήγορα και εύκολα, αυτή η μικροσκοπική μύγα χρησιμεύει ως πρότυπο οργανισμό για τους εξελικτικούς βιολόγους. Το χρησιμοποιούν για να ρίξουν φως στο θαύμα της δημιουργίας - από τη στιγμή της γονιμοποίησης μέχρι την εμφάνιση ενός ενήλικου οργανισμού. Χάρη στη Drosophila, έχουν γίνει πολλές ανακαλύψεις, συμπεριλαμβανομένης της ανακάλυψης γονιδίων που περιέχουν homeobox που ρυθμίζουν τη γενική δομή όλων των ζωντανών οργανισμών.
Κάθε σπουδαστής γενετικής είναι εξοικειωμένος με τα πειράματα στη Drosophila που έκανε ο Thomas Hunt Morgan, ο πατέρας της αμερικανικής γενετικής. Το 1910, παρατήρησε αρσενικούς μεταλλαγμένους με λευκά μάτια ανάμεσα στις συνηθισμένες μύγες με κόκκινα μάτια. Διασταύρωσε ένα αρσενικό με λευκά μάτια με ένα θηλυκό με κόκκινα μάτια και διαπίστωσε ότι οι απόγονοί τους ήταν κόκκινα μάτια: η ασπρομάτα αποδείχθηκε ότι ήταν ένα υπολειπόμενο χαρακτηριστικό και τώρα γνωρίζουμε ότι για να έχουν λευκά μάτια οι μύγες, χρειάζονται δύο αντίγραφα του γονιδίου των λευκών ματιών, ένα από κάθε γονέα. Συνεχίζοντας να διασταυρώνει μεταλλαγμένα, ο Morgan ανακάλυψε ότι μόνο τα αρσενικά εμφάνιζαν το χαρακτηριστικό των λευκών ματιών και κατέληξε στο συμπέρασμα ότι αυτό το χαρακτηριστικό συσχετίστηκε με το φυλετικό χρωμόσωμα (χρωμόσωμα Υ). Ο Μόργκαν και οι μαθητές του μελέτησαν κληρονομικά χαρακτηριστικά σε χιλιάδες μύγες φρούτων. Σήμερα, πειράματα με τη Drosophila πραγματοποιούνται σε εργαστήρια μοριακής βιολογίας σε όλο τον κόσμο, όπου περισσότεροι από πέντε χιλιάδες άνθρωποι μελετούν αυτό το μικρό έντομο.
Έμαθα από πρώτο χέρι τη σημασία του Drosophila όταν χρησιμοποίησα βιβλιοθήκες των γονιδίων του cDNA για να μελετήσω τους υποδοχείς αδρεναλίνης και ανακάλυψα το ισοδύναμό τους στους υποδοχείς της μύγας - οκτοπαμίνης. Αυτή η ανακάλυψη έδειξε την κοινότητα της εξελικτικής κληρονομικότητας του νευρικού συστήματος της μύγας και του ανθρώπου. Προσπαθώντας να κατανοήσω τις βιβλιοθήκες cDNA του ανθρώπινου εγκεφάλου, βρήκα γονίδια με παρόμοιες λειτουργίες με σύγκριση ανθρώπινων γονιδίων με υπολογιστή με γονίδια Drosophila.
Το έργο προσδιορισμού αλληλουχίας γονιδίων Drosophila ξεκίνησε το 1991, όταν ο Jerry Rubin από το Πανεπιστήμιο της Καλιφόρνια στο Μπέρκλεϋ και ο Allen Spradling του Ινστιτούτου Carnegie αποφάσισαν ότι ήταν καιρός να αναλάβουν το έργο. Μέχρι τον Μάιο του 1998, το 25% της αλληλουχίας είχε ήδη ολοκληρωθεί και έκανα μια πρόταση που ο Ρούμπιν είπε ότι ήταν «πολύ καλή για να την αφήσω». Η ιδέα μου ήταν αρκετά επικίνδυνη: χιλιάδες ερευνητές μυγών από διαφορετικές χώρες θα έπρεπε να εξετάσουν προσεκτικά κάθε γράμμα του κώδικα που λάβαμε, συγκρίνοντάς το με τα υψηλής ποιότητας, δεδομένα αναφοράς του ίδιου του Jerry, και στη συνέχεια να βγάλουν ένα συμπέρασμα σχετικά με την καταλληλότητα της μεθόδου μου .
Το αρχικό σχέδιο ήταν να ολοκληρωθεί η αλληλουχία του γονιδιώματος της μύγας εντός έξι μηνών έως τον Απρίλιο του 1999 και στη συνέχεια να ξεκινήσει η επίθεση στο ανθρώπινο γονιδίωμα. Μου φάνηκε ότι αυτός ήταν ο πιο αποτελεσματικός και σαφής τρόπος για να δείξουμε ότι η νέα μας μέθοδος λειτουργεί. Και αν δεν τα καταφέρουμε, σκέφτηκα, τότε θα ήταν καλύτερο να το επαληθεύσουμε γρήγορα χρησιμοποιώντας το παράδειγμα της Drosophila παρά δουλεύοντας το ανθρώπινο γονιδίωμα. Αλλά στην πραγματικότητα, η πλήρης αποτυχία θα ήταν η πιο θεαματική αποτυχία στην ιστορία της βιολογίας. Ο Τζέρι έβαζε επίσης τη φήμη του στη γραμμή, οπότε όλοι στο Celera ήταν αποφασισμένοι να τον υποστηρίξουν. Ζήτησα από τον Mark Adams να ηγηθεί του μέρους μας στο έργο, και επειδή ο Jerry είχε επίσης μια κορυφαία ομάδα στο Berkeley, η συνεργασία μας πήγε καλά.
Πρώτα απ 'όλα, προέκυψε το ερώτημα σχετικά με την καθαρότητα του DNA που έπρεπε να ακολουθήσουμε. Όπως οι άνθρωποι, οι μύγες ποικίλλουν σε γενετικό επίπεδο. Εάν υπάρχει πάνω από 2% γενετική διαφοροποίηση σε έναν πληθυσμό και έχουμε 50 διαφορετικά άτομα στην επιλεγμένη ομάδα, τότε η αποκωδικοποίηση αποδεικνύεται πολύ δύσκολη. Το πρώτο βήμα του Τζέρι ήταν να ενσωματώσει τις μύγες όσο το δυνατόν περισσότερο για να μας δώσει μια ομοιόμορφη παραλλαγή DNA. Αλλά η ενδογαμία δεν ήταν αρκετή για να διασφαλίσει τη γενετική καθαρότητα: κατά την εξαγωγή DNA μύγας, υπήρχε κίνδυνος μόλυνσης με γενετικό υλικό από βακτηριακά κύτταρα στην τροφή της μύγας ή στα έντερα της. Για να αποφύγει αυτά τα προβλήματα, ο Τζέρι προτίμησε να εξάγει DNA από έμβρυα μύγας. Αλλά ακόμη και από εμβρυϊκά κύτταρα, έπρεπε πρώτα να απομονώσουμε πυρήνες με το DNA που χρειαζόμασταν, ώστε να μην το μολύνουμε με εξωπυρηνικό DNA των μιτοχονδρίων - τα «ηλεκτρικά εργοστάσια» του κυττάρου. Ως αποτέλεσμα, λάβαμε έναν δοκιμαστικό σωλήνα με ένα θολό διάλυμα καθαρού DNA Drosophila.
Το καλοκαίρι του 1998, η ομάδα του Ham, έχοντας τόσο καθαρό DNA μύγας, άρχισε να δημιουργεί βιβλιοθήκες με τα θραύσματά του. Ο ίδιος ο Ham άρεσε περισσότερο να κόβει DNA και να επικαλύπτει τα θραύσματα που προκύπτουν, μειώνοντας την ευαισθησία του ακουστικού του, έτσι ώστε κανένας ξένος ήχος να μην τον αποσπά την προσοχή από τη δουλειά του. Η δημιουργία βιβλιοθηκών υποτίθεται ότι ήταν η αρχή μιας μεγάλης κλίμακας αλληλουχίας, αλλά μέχρι στιγμής μόνο οι ήχοι από τρυπάνια, σφυριά και πριόνια ακούγονταν παντού. Ένας ολόκληρος στρατός κατασκευαστών βρισκόταν συνεχώς στο μάτι κοντά και συνεχίσαμε να λύνουμε τα πιο σημαντικά προβλήματα - αντιμετώπιση προβλημάτων στη λειτουργία των sequencers, ρομπότ και άλλου εξοπλισμού, προσπαθώντας όχι σε χρόνια, αλλά σε λίγους μήνες να δημιουργήσουμε ένα πραγματικό «εργοστάσιο» αλληλουχίας " από την αρχή.
Ο πρώτος προσδιοριστής αλληλουχίας DNA Model 3700 παραδόθηκε στη Celera στις 8 Δεκεμβρίου 1998 με μεγάλο ενθουσιασμό και συλλογικό αναστεναγμό ανακούφισης. Η συσκευή αφαιρέθηκε από ένα ξύλινο κουτί, τοποθετήθηκε σε ένα δωμάτιο χωρίς παράθυρα στο υπόγειο - το προσωρινό της σπίτι, και άρχισε αμέσως η δοκιμή. Μόλις άρχισε να λειτουργεί, είχαμε αποτελέσματα πολύ υψηλής ποιότητας. Αλλά αυτά τα πρώιμα sequencer ήταν αρκετά ασταθή και μερικά ήταν ελαττωματικά από την αρχή. Υπήρχαν επίσης συνεχή προβλήματα με τους εργάτες, μερικές φορές σχεδόν καθημερινά. Για παράδειγμα, εμφανίστηκε ένα σοβαρό σφάλμα στο πρόγραμμα ελέγχου του ρομποτικού χειριστή - μερικές φορές ο μηχανικός βραχίονας του ρομπότ εκτεινόταν πάνω από τη συσκευή με υψηλή ταχύτητα και έπεσε στον τοίχο. Ως αποτέλεσμα, το sequencer σταμάτησε και έπρεπε να κληθεί μια ομάδα επισκευής για να το διορθώσει. Ορισμένα συστήματα αλληλουχίας απέτυχαν λόγω αδέσποτων ακτίνων λέιζερ. Για την προστασία από την υπερθέρμανση, χρησιμοποιήθηκαν αλουμινόχαρτο και ταινία, καθώς σε υψηλές θερμοκρασίες τα κίτρινα θραύσματα Gs εξατμίστηκαν από τις αλληλουχίες.
Παρόλο που οι συσκευές προμηθεύονταν πλέον τακτικά, περίπου το 90% από αυτές ήταν ελαττωματικές από την αρχή. Κάποιες μέρες τα sequencer δεν λειτουργούσαν καθόλου. Πίστευα ακράδαντα στον Mike Hunkapiller, αλλά η πίστη μου κλονίστηκε πολύ όταν άρχισε να κατηγορεί τις αποτυχίες μας στους υπαλλήλους μας, τη σκόνη των κατασκευών, τις παραμικρές διακυμάνσεις της θερμοκρασίας, τις φάσεις του φεγγαριού κ.λπ. Κάποιοι από εμάς έγιναν γκρι από το άγχος.
Οι νεκροί 3700 κάθονταν στην καφετέρια και περίμεναν να τους στείλουν πίσω στο ABI, και τελικά έφτασε στο σημείο που έπρεπε να φάμε μεσημεριανό σχεδόν στο νεκροτομείο των sequencers. Ήμουν σε απόγνωση - τελικά, χρειαζόμουν έναν συγκεκριμένο αριθμό συσκευών που λειτουργούσαν κάθε μέρα, δηλαδή 230! Για περίπου 70 εκατομμύρια δολάρια, η ABI υποσχέθηκε να μας παράσχει είτε 230 τέλεια λειτουργικές συσκευές που θα λειτουργούσαν όλη την ημέρα χωρίς διακοπή, είτε 460 που θα λειτουργούσαν για τουλάχιστον μισή μέρα. Επιπλέον, ο Mike θα έπρεπε να έχει διπλασιάσει τον αριθμό του ειδικευμένου τεχνικού προσωπικού για την άμεση επισκευή των sequencers μετά από αστοχία.
Τι συμφέρον όμως να τα κάνεις όλα αυτά στα ίδια λεφτά! Επιπλέον, ο Mike έχει τώρα έναν άλλο πελάτη - ένα κυβερνητικό έργο γονιδιώματος, του οποίου οι ηγέτες έχουν ήδη αρχίσει να αγοράζουν εκατοντάδες συσκευές χωρίς καμία δοκιμή. Το μέλλον του Celera εξαρτιόταν από αυτά τα sequencers, αλλά ο Mike προφανώς δεν συνειδητοποίησε ότι το μέλλον του ABI εξαρτιόταν επίσης από αυτούς. Η σύγκρουση ήταν αναπόφευκτη, όπως φάνηκε σε μια σημαντική συνάντηση μεταξύ των μηχανικών της ABI και της ομάδας μου στη Celera.
Αφού αναφέραμε τον τεράστιο αριθμό ελαττωματικών οργάνων και τον χρόνο που χρειάστηκε για να διορθωθούν οι βλάβες του sequencer, ο Mike προσπάθησε και πάλι να ρίξει όλη την ευθύνη στους υπαλλήλους μου, αλλά ακόμη και οι δικοί του μηχανικοί διαφώνησαν μαζί του. Ο Τόνι Γουάιτ τελικά παρενέβη. «Δεν με νοιάζει πόσο κοστίζει ή ποιος πρέπει να σκοτωθεί για αυτό», είπε. Τότε για πρώτη και τελευταία φορά πήρε πραγματικά το μέρος μου. Διέταξε τον Mike να διασφαλίσει την παράδοση των νέων sequencer όσο το δυνατόν γρηγορότερα, ακόμη και σε βάρος άλλων πελατών και ακόμη και αν δεν ήταν ακόμη γνωστό πόσο θα κόστιζε.
Ο Τόνι διέταξε επίσης τον Μάικ να προσλάβει είκοσι ακόμη τεχνικούς για να επισκευάσουν γρήγορα και να προσδιορίσουν την αιτία όλων των προβλημάτων. Στην πραγματικότητα, αυτό ήταν πιο εύκολο να ειπωθεί παρά να γίνει, επειδή οι έμπειροι εργαζόμενοι ήταν σε έλλειψη. Αρχικά, ο Έρικ Λάντερ έκανε λαθροθηρία δύο από τους πιο καταρτισμένους μηχανικούς και κατά τη γνώμη του Μάικ, φταίμε και εμείς. Γυρίζοντας στον Μαρκ Άνταμς, ο Μάικ είπε: «Έπρεπε να τους είχες προσλάβει πριν το κάνει κάποιος άλλος». Μετά από μια τέτοια δήλωση, έχασα εντελώς κάθε σεβασμό για αυτόν. Εξάλλου, σύμφωνα με τη συμφωνία μας, δεν μπορούσα να προσλάβω υπαλλήλους της ABI, ενώ ο Lander και άλλοι ηγέτες του κυβερνητικού προγράμματος γονιδιώματος είχαν το δικαίωμα να το κάνουν, οπότε πολύ σύντομα οι καλύτεροι μηχανικοί της ABI άρχισαν να εργάζονται για τους ανταγωνιστές μας. Μέχρι το τέλος της συνάντησης, συνειδητοποίησα ότι τα προβλήματα παρέμεναν, αλλά μια αχτίδα ελπίδας για βελτίωση είχε ανατείλει.
Και έτσι έγινε, αν και όχι αμέσως. Το οπλοστάσιο των sequencers μας αυξήθηκε από 230 σε 300 συσκευές, και αν το 20-25% από αυτά αποτύγχανε, είχαμε ακόμα περίπου 200 εργάσιμους sequencer και με κάποιο τρόπο ανταπεξέλθαμε στις εργασίες. Το τεχνικό επιτελείο εργάστηκε ηρωικά και ανέβαζε σταθερά τους ρυθμούς των εργασιών επισκευής, μειώνοντας το χρόνο διακοπής λειτουργίας. Όλο αυτό το διάστημα σκεφτόμουν ένα πράγμα: αυτό που κάνουμε είναι εφικτό. Οι αποτυχίες συνέβησαν για χίλιους λόγους, αλλά η αποτυχία δεν ήταν μέρος των σχεδίων μου.
Ξεκινήσαμε σοβαρά τον προσδιορισμό της αλληλουχίας του γονιδιώματος της Drosophila στις 8 Απριλίου, περίπου τη στιγμή που θα έπρεπε να είχαμε ολοκληρώσει αυτήν την εργασία. Φυσικά, κατάλαβα ότι ο White ήθελε να με ξεφορτωθεί, αλλά έκανα ό,τι περνούσε από το χέρι μου για να ολοκληρώσω το κύριο έργο. Ένταση και άγχος με στοίχειωναν στο σπίτι, αλλά δεν μπορούσα να συζητήσω αυτά τα προβλήματα με τον «έμπιστο» μου. Η Κλερ έδειξε την περιφρόνησή της όταν είδε πόσο με απασχολούσαν οι υποθέσεις της Σέλαρα. Ένιωθε σαν να επαναλάμβανα τα ίδια λάθη που έκανα ενώ δούλευα στο TIGR/HGS. Την 1η Ιουλίου ένιωθα βαθιά κατάθλιψη, όπως και στο Βιετνάμ.
Δεδομένου ότι η μέθοδος του μεταφορέα δεν είχε ακόμη λειτουργήσει για εμάς, έπρεπε να κάνουμε σκληρή, εξαντλητική δουλειά - να «κολλήσουμε» τα θραύσματα του γονιδιώματος ξανά μεταξύ τους. Για να ανιχνεύσει αγώνες χωρίς να αποσπάται η προσοχή από επαναλήψεις, ο Gene Myers πρότεινε έναν αλγόριθμο που βασίζεται στη βασική αρχή της δικής μου εκδοχής της μεθόδου του κυνηγετικού όπλου: ακολουθήστε και τα δύο άκρα όλων των κλώνων που προέκυψαν. Επειδή ο Ham έπαιρνε κλώνους τριών επακριβώς γνωστών μεγεθών, γνωρίζαμε ότι οι δύο τερματικές ακολουθίες βρίσκονταν σε μια αυστηρά καθορισμένη απόσταση μεταξύ τους. Όπως και πριν, αυτή η μέθοδος «ταιριάσματος» θα μας δώσει μια εξαιρετική ευκαιρία να επανασυναρμολογήσουμε το γονιδίωμα.
Αλλά επειδή κάθε άκρο της ακολουθίας αλληλουχήθηκε ξεχωριστά, για να λειτουργήσει με ακρίβεια αυτή η μέθοδος συναρμολόγησης, ήταν απαραίτητο να τηρούνται προσεκτικά αρχεία - για να είμαστε απολύτως βέβαιοι ότι μπορούσαμε να συνδέσουμε σωστά όλα τα ζεύγη τελικών ακολουθιών: τελικά, αν τουλάχιστον μία στις εκατό προσπάθειες οδηγεί σε σφάλμα και δεν βρέθηκε καμία αντίστοιχη, ένα ζευγάρι για συνέπεια, όλα θα πάνε κάτω και η μέθοδος δεν θα λειτουργήσει. Ένας τρόπος για να αποφευχθεί αυτό είναι να χρησιμοποιήσετε γραμμωτούς κώδικες και αισθητήρες για να παρακολουθείτε κάθε βήμα της διαδικασίας. Αλλά στην αρχή της εργασίας, οι τεχνικοί εργαστηρίου δεν είχαν το απαραίτητο λογισμικό και εξοπλισμό για την αλληλουχία, έτσι έπρεπε να κάνουν τα πάντα χειροκίνητα. Στο Celera, μια μικρή ομάδα λιγότερων από είκοσι ατόμων επεξεργαζόταν ένα ρεκόρ 200.000 κλώνων κάθε μέρα. Θα μπορούσαμε να προβλέψουμε ορισμένα σφάλματα, όπως η εσφαλμένη ανάγνωση δεδομένων από 384 πηγαδάκια και, στη συνέχεια, να χρησιμοποιήσουμε τον υπολογιστή για να εντοπίσουμε την εμφανώς λανθασμένη λειτουργία και να διορθώσουμε την κατάσταση. Φυσικά, υπήρχαν ακόμα κάποιες ελλείψεις, αλλά αυτό επιβεβαίωσε μόνο την ικανότητα και τη σιγουριά της ομάδας ότι μπορούσαμε να εξαλείψουμε τα λάθη.
Παρά όλες τις δυσκολίες, μπορέσαμε να διαβάσουμε 3156 εκατομμύρια αλληλουχίες σε τέσσερις μήνες, συνολικά περίπου 1,76 δισεκατομμύρια ζεύγη νουκλεοτιδίων που περιέχονται μεταξύ των άκρων 1,51 εκατομμυρίων κλώνων DNA. Τώρα ήταν η σειρά του Gene Myers, της ομάδας του και του υπολογιστή μας - ήταν απαραίτητο να ενωθούν όλα τα τμήματα μαζί στα χρωμοσώματα Drosophila. Όσο μακρύτερες γίνονταν οι τομές, τόσο λιγότερο ακριβής γινόταν η αλληλουχία. Στην περίπτωση της Drosophila, οι αλληλουχίες ήταν κατά μέσο όρο 551 ζεύγη βάσεων και η μέση ακρίβεια ήταν 99,5%. Λαμβάνοντας υπόψη τις ακολουθίες 500 γραμμάτων, σχεδόν ο καθένας μπορεί να εντοπίσει τους αγώνες μετακινώντας τη μια ακολουθία κατά μήκος μιας άλλης μέχρι να βρεθεί ένα ταίριασμα.
Για την αλληλουχία του Haemophilus influenzae, είχαμε 26 χιλιάδες αλληλουχίες. Για να συγκριθεί το καθένα από αυτά με όλα τα άλλα απαιτούνται 26 χιλιάδες συγκρίσεις στο τετράγωνο, ή 676 εκατομμύρια. Το γονιδίωμα της Drosophila, με τις 3,156 εκατομμύρια αναγνώσεις, θα απαιτούσε περίπου 9,9 τρισεκατομμύρια συγκρίσεις. Στην περίπτωση του ανθρώπου και του ποντικιού, όπου δημιουργήσαμε 26 εκατομμύρια αναγνώσεις ακολουθίας, απαιτήθηκαν περίπου 680 τρισεκατομμύρια συγκρίσεις. Επομένως, δεν προκαλεί έκπληξη το γεγονός ότι οι περισσότεροι επιστήμονες ήταν πολύ δύσπιστοι σχετικά με την πιθανή επιτυχία αυτής της μεθόδου.
Αν και ο Myers υποσχέθηκε να τα φτιάξει όλα, είχε συνεχώς αμφιβολίες. Τώρα δούλευε μέρες και νύχτες, φαινόταν εξαντλημένος και κάπως γκρίζος. Επιπλέον, είχε προβλήματα στην οικογένειά του και άρχισε να περνά τον περισσότερο ελεύθερο χρόνο του με τον δημοσιογράφο James Shreve, ο οποίος έγραψε για το έργο μας και, σαν σκιά, παρακολουθούσε την πρόοδο της έρευνας. Προσπαθώντας με κάποιο τρόπο να αποσπάσω την προσοχή του Τζιν, τον πήρα μαζί μου στην Καραϊβική για να χαλαρώσω και να πλεύσω με το γιοτ μου. Αλλά ακόμα κι εκεί καθόταν για ώρες, σκυμμένος πάνω από το λάπτοπ του, συνοφρυώνοντας τα μαύρα φρύδια του και στραβίζοντας τα μαύρα μάτια του από τον λαμπερό ήλιο. Και, παρά τις απίστευτες δυσκολίες, ο Gene και η ομάδα του κατάφεραν να δημιουργήσουν περισσότερες από μισό εκατομμύριο γραμμές κώδικα υπολογιστή για το νέο assembler σε έξι μήνες.
Εάν τα αποτελέσματα της αλληλουχίας ήταν 100% ακριβή, χωρίς διπλό DNA, η συναρμολόγηση του γονιδιώματος θα ήταν μια σχετικά απλή εργασία. Αλλά στην πραγματικότητα, τα γονιδιώματα περιέχουν μεγάλο αριθμό επαναλαμβανόμενων DNA διαφορετικών τύπων, μηκών και συχνοτήτων. Οι σύντομες επαναλήψεις με λιγότερα από πεντακόσια ζεύγη βάσεων είναι σχετικά εύκολο να αντιμετωπιστούν· οι μεγαλύτερες επαναλήψεις είναι πιο δύσκολες. Για να λύσουμε αυτό το πρόβλημα, χρησιμοποιήσαμε μια μέθοδο «εύρεσης ζεύγους», δηλαδή προσδιορίσαμε την αλληλουχία και των δύο άκρων κάθε κλώνου και λάβαμε κλώνους διαφορετικού μήκους για να εξασφαλίσουμε τον μέγιστο αριθμό αντιστοιχιών.
Οι αλγόριθμοι, κωδικοποιημένοι στις μισές γραμμές κώδικα υπολογιστή της ομάδας του Jin, πρότειναν ένα σενάριο βήμα προς βήμα - από τις πιο «ακίνδυνες» ενέργειες, όπως η απλή επικάλυψη δύο ακολουθιών, έως πιο περίπλοκες, όπως η χρήση ανιχνευμένων ζευγών για συγχώνευση νησίδων αλληλοκαλυπτόμενων ακολουθιών. Ήταν σαν να φτιάχνεις ένα παζλ, όπου μικρές νησίδες συναρμολογημένων τμημάτων ενώνονται για να σχηματίσουν μεγαλύτερα νησιά και μετά η όλη διαδικασία επαναλαμβάνεται ξανά. Μόνο το παζλ μας είχε 27 εκατομμύρια κομμάτια. Και ήταν πολύ σημαντικό ότι τα τμήματα είχαν ληφθεί από μια ακολουθία συναρμολόγησης υψηλής ποιότητας: φανταστείτε τι θα συνέβαινε αν συναρμολογούσατε ένα παζλ και τα χρώματα ή οι εικόνες των στοιχείων του είναι ασαφή και θολά. Για σειρά μακράς εμβέλειας της αλληλουχίας του γονιδιώματος, ένα σημαντικό ποσοστό των αναγνώσεων πρέπει να έχει τη μορφή ταιριασμένων ζευγών. Δεδομένου ότι τα αποτελέσματα εξακολουθούσαν να παρακολουθούνται χειροκίνητα, ανακουφιστήκαμε όταν διαπιστώσαμε ότι το 70% των ακολουθιών που είχαμε ήταν ακριβώς έτσι. Οι μοντελιστές υπολογιστών εξήγησαν ότι με χαμηλότερο ποσοστό θα ήταν αδύνατο να συναρμολογήσουμε το "Humpty Dumpty".
Και τώρα μπορέσαμε να χρησιμοποιήσουμε το assembler Celera για να ακολουθήσουμε την ακολουθία: στο πρώτο στάδιο, τα αποτελέσματα προσαρμόστηκαν για να επιτευχθεί η υψηλότερη ακρίβεια. στο δεύτερο βήμα, ο Screener αφαίρεσε τις μολυσματικές αλληλουχίες από το πλασμίδιο ή το DNA του Ε. coli. Η διαδικασία συναρμολόγησης μπορεί να διαταραχθεί από μόλις 10 ζεύγη βάσεων της «ξένης» ακολουθίας. Στο τρίτο βήμα, το πρόγραμμα Screener έλεγξε κάθε θραύσμα για συμμόρφωση με γνωστές επαναλαμβανόμενες αλληλουχίες στο γονιδίωμα της μύγας των φρούτων - δεδομένα από τον Jerry Rubin, ο οποίος μας τα παρείχε «ευγενικά». Καταγράφηκαν οι θέσεις των επαναλήψεων με μερικώς επικαλυπτόμενες περιοχές. Στο τέταρτο βήμα, ένα άλλο πρόγραμμα (Overlapper) ανακάλυψε τις επικαλυπτόμενες περιοχές συγκρίνοντας κάθε θραύσμα με όλα τα άλλα - ένα κολοσσιαίο πείραμα στην επεξεργασία ενός τεράστιου όγκου αριθμητικών δεδομένων. Συγκρίναμε 32 εκατομμύρια θραύσματα κάθε δευτερόλεπτο, με στόχο να βρούμε τουλάχιστον 40 επικαλυπτόμενα ζεύγη βάσεων με λιγότερο από 6% διαφορές. Όταν ανακαλύψαμε δύο επικαλυπτόμενες περιοχές, τις συνδυάσαμε σε ένα μεγαλύτερο θραύσμα, το λεγόμενο "contig" - ένα σύνολο επικαλυπτόμενων θραυσμάτων.
Στην ιδανική περίπτωση, αυτό θα ήταν αρκετό για τη συναρμολόγηση του γονιδιώματος. Αλλά έπρεπε να αντιμετωπίσουμε τα τραύλισμα και τις επαναλήψεις στον κώδικα του DNA, πράγμα που σήμαινε ότι ένα κομμάτι DNA θα μπορούσε να επικαλύπτεται με πολλές διαφορετικές περιοχές, δημιουργώντας ψευδείς συνδέσεις. Για να απλοποιήσουμε την εργασία, αφήσαμε μόνο μοναδικά συνδεδεμένα θραύσματα, τα λεγόμενα "unitigs". Το πρόγραμμα που χρησιμοποιήσαμε για να εκτελέσουμε αυτή τη λειτουργία (Unitigger) ουσιαστικά αφαίρεσε όλη την αλληλουχία DNA που δεν μπορούσαμε να αναγνωρίσουμε με βεβαιότητα, αφήνοντας μόνο αυτές τις μονάδες. Αυτό το βήμα όχι μόνο μας έδωσε την ευκαιρία να εξετάσουμε άλλες επιλογές για τη συναρμολόγηση των θραυσμάτων, αλλά και απλοποίησε σημαντικά την εργασία. Μετά τη μείωση, ο αριθμός των επικαλυπτόμενων θραυσμάτων μειώθηκε από 212 εκατομμύρια σε 3,1 εκατομμύρια και το πρόβλημα απλοποιήθηκε κατά 68 φορές. Τα κομμάτια του παζλ σταδιακά αλλά σταθερά μπήκαν στη θέση τους.
Και τότε θα μπορούσαμε να χρησιμοποιήσουμε πληροφορίες σχετικά με τον τρόπο που οι ακολουθίες του ίδιου κλώνου ζευγαρώθηκαν χρησιμοποιώντας έναν αλγόριθμο «σκελετού». Όλες οι πιθανές μονάδες με αμοιβαία επικαλυπτόμενα ζεύγη βάσεων συνδυάστηκαν σε ειδικά πλαίσια. Για να περιγράψω αυτό το στάδιο στις διαλέξεις μου, σχεδιάζω μια αναλογία με το σετ κατασκευής παιδικών παιχνιδιών Tinkertoys. Αποτελείται από ξυλάκια διαφορετικού μήκους, τα οποία μπορούν να εισαχθούν σε τρύπες που βρίσκονται σε ξύλινα βασικά μέρη (μπάλες και δίσκοι), και έτσι να δημιουργήσουν μια τρισδιάστατη δομή. Στην περίπτωσή μας, τα βασικά μέρη είναι μονάδες. Γνωρίζοντας ότι οι ζευγαρωμένες αλληλουχίες βρίσκονται στα άκρα των κλώνων μήκους 2 χιλιάδων, 10 χιλιάδων ή 50 χιλιάδων ζευγών βάσεων - δηλαδή, φαίνονται να βρίσκονται σε απόσταση ενός συγκεκριμένου αριθμού οπών η μία από την άλλη - μπορούν να παραταχθούν.
Η δοκιμή αυτής της τεχνικής στην αλληλουχία του Jerry Rubin, η οποία ήταν περίπου το ένα πέμπτο του γονιδιώματος της μύγας των φρούτων, είχε ως αποτέλεσμα μόνο 500 κενά. Δοκιμάζοντας τα δικά μας δεδομένα τον Αύγουστο, καταλήξαμε με περισσότερα από 800.000 μικρά θραύσματα. Ένας σημαντικά μεγαλύτερος όγκος δεδομένων για επεξεργασία έδειξε ότι η τεχνική λειτούργησε άσχημα - το αποτέλεσμα ήταν το αντίθετο από αυτό που αναμενόταν. Τις επόμενες μέρες, ο πανικός μεγάλωσε και η λίστα με τα πιθανά λάθη επεκτάθηκε. Από τον τελευταίο όροφο του κτιρίου Νο. 2, μια έκρηξη αδρεναλίνης μπήκε στο δωμάτιο που ονομαζόταν χαριτολογώντας «Ήσυχα δωμάτια». Ωστόσο, δεν υπήρχε αίσθηση γαλήνης ή γαλήνης εκεί, ειδικά για τουλάχιστον μια-δυο εβδομάδες, όταν οι εργαζόμενοι κυριολεκτικά τριγυρνούσαν σε κύκλους, αναζητώντας μια διέξοδο από την κατάσταση.
Το πρόβλημα τελικά λύθηκε από τον Arthur Delcher, ο οποίος εργάστηκε με το πρόγραμμα Overlapper. Παρατήρησε κάτι περίεργο στη γραμμή 678 από τις 150.000 γραμμές κώδικα, όπου μια μικρή απόκλιση σήμαινε ότι ένα σημαντικό μέρος του αγώνα δεν καταγράφηκε. Το σφάλμα διορθώθηκε και στις 7 Σεπτεμβρίου είχαμε 134 κυτταρικά ικριώματα που κάλυπταν το πραγματικό (ευχρωματικό) γονιδίωμα της μύγας των φρούτων. Ήμασταν ενθουσιασμένοι και πήραμε έναν αναστεναγμό ανακούφισης. Ήρθε η ώρα να ανακοινώσουμε την επιτυχία μας σε όλο τον κόσμο.
Το συνέδριο για την αλληλουχία γονιδιώματος, το οποίο ξεκίνησα να φιλοξενώ πριν από αρκετά χρόνια, έδωσε μια εξαιρετική ευκαιρία για αυτό. Ήμουν σίγουρος ότι θα υπήρχε ένας μεγάλος αριθμός ανθρώπων που θα ήθελαν να βεβαιωθούν αν είχαμε κρατήσει την υπόσχεσή μας. Αποφάσισα ότι ο Mark Adams, ο Gene Myers και ο Jerry Rubin θα πρέπει να μιλήσουν για τα επιτεύγματά μας, και κυρίως για τη διαδικασία αλληλούχισης, τη συναρμολόγηση του γονιδιώματος και τη σημασία αυτού για την επιστήμη. Λόγω της εισροής ανθρώπων που ήθελαν να έρθουν στο συνέδριο, έπρεπε να το μεταφέρω από το Hilton Head στο μεγαλύτερο ξενοδοχείο Fontainebleau στο Μαϊάμι. Στο συνέδριο συμμετείχαν εκπρόσωποι μεγάλων φαρμακευτικών και βιοτεχνολογικών εταιρειών, ειδικοί στη γονιδιωματική έρευνα από όλο τον κόσμο, αρκετοί αρθρογράφοι, δημοσιογράφοι και εκπρόσωποι επενδυτικών εταιρειών - όλοι ήταν εκεί. Οι ανταγωνιστές μας από το Incyte ξόδεψαν πολλά χρήματα για τη διοργάνωση μιας δεξίωσης μετά το συνέδριο, εταιρική βιντεοσκόπηση κ.λπ. - έκαναν τα πάντα για να πείσουν το κοινό ότι πρόσφεραν «τις πιο λεπτομερείς πληροφορίες για το ανθρώπινο γονιδίωμα».
Μαζευτήκαμε σε μια μεγάλη αίθουσα συνεδριάσεων. Διακοσμημένο σε ουδέτερα χρώματα, διακοσμημένο με φωτιστικά τοίχου, σχεδιάστηκε για δύο χιλιάδες άτομα, αλλά ο κόσμος συνέχιζε να έρχεται και σύντομα η αίθουσα γέμισε στο έπακρο. Το συνέδριο ξεκίνησε στις 17 Σεπτεμβρίου 1999, με παρουσιάσεις από τους Jerry, Mark και Gene στην πρώτη συνεδρία. Μετά από μια σύντομη εισαγωγή, ο Jerry Rubin ανακοίνωσε ότι το κοινό επρόκειτο να ακούσει για το καλύτερο κοινό έργο διάσημων εταιρειών στις οποίες είχε εμπλακεί ποτέ. Η ατμόσφαιρα ζεσταινόταν. Το κοινό συνειδητοποίησε ότι δεν θα μιλούσε τόσο πομπωδώς αν δεν είχαμε ετοιμάσει κάτι πραγματικά συγκλονιστικό.
Στη σιωπή που ακολούθησε, ο Mark Adams άρχισε να περιγράφει λεπτομερώς τη δουλειά του «κατασκευασμένου καταστήματος» μας στη Celera και τις νέες μας μεθόδους προσδιορισμού αλληλουχίας γονιδιώματος. Ωστόσο, δεν είπε λέξη για το συναρμολογημένο γονιδίωμα, σαν να πείραζε το κοινό. Στη συνέχεια ο Gene βγήκε και μίλησε για τις αρχές της μεθόδου του κυνηγετικού όπλου, για την αλληλουχία του Haemophilus και για τα κύρια στάδια του assembler. Χρησιμοποιώντας κινούμενα σχέδια υπολογιστή, έδειξε ολόκληρη τη διαδικασία της αντίστροφης συναρμολόγησης του γονιδιώματος. Ο χρόνος που είχε διατεθεί για τις παρουσιάσεις τελείωνε και πολλοί είχαν ήδη αποφασίσει ότι όλα θα περιορίζονταν σε μια στοιχειώδη παρουσίαση χρησιμοποιώντας το PowerPoint, χωρίς να παρουσιάζονται συγκεκριμένα αποτελέσματα. Αλλά στη συνέχεια ο Gene σημείωσε με ένα κακόβουλο χαμόγελο ότι το κοινό πιθανότατα θα ήθελε ακόμα να δει πραγματικά αποτελέσματα και δεν θα αρκέστηκε σε μια μίμηση.
Ήταν αδύνατο να παρουσιάσουμε τα αποτελέσματά μας πιο ξεκάθαρα και εκφραστικά από ό,τι έκανε ο Gene Myers. Συνειδητοποίησε ότι τα αποτελέσματα της αλληλουχίας από μόνα τους δεν θα έκαναν τη σωστή εντύπωση, οπότε για να γίνει πιο πειστικό, τα συνέκρινε με τα αποτελέσματα της επίπονης έρευνας του Τζέρι χρησιμοποιώντας την παραδοσιακή μέθοδο. Αποδείχτηκαν πανομοιότυποι! Έτσι, ο Jin συνέκρινε τα αποτελέσματα της συναρμολόγησης του γονιδιώματος μας με όλους τους γνωστούς δείκτες που είχαν χαρτογραφηθεί στο γονιδίωμα της μύγας φρούτων πριν από δεκαετίες. Από χιλιάδες δείκτες, μόνο έξι δεν ταιριάζουν με τα αποτελέσματα της συνέλευσής μας. Εξετάζοντας προσεκτικά και τα έξι, ήμασταν πεπεισμένοι ότι η αλληλουχία του Celera ήταν σωστή και ότι περιέχονταν σφάλματα σε εργασίες που έγιναν σε άλλα εργαστήρια χρησιμοποιώντας παλιές μεθόδους. Τέλος, ο Gene είπε ότι μόλις ξεκινήσαμε τον προσδιορισμό της αλληλουχίας του ανθρώπινου DNA και οι επαναλήψεις θα ήταν πιθανώς λιγότερο πρόβλημα από ό,τι με τη Drosophila.
Ακολούθησαν δυνατά και παρατεταμένα χειροκροτήματα. Ο βρυχηθμός που δεν σταμάτησε στο διάλειμμα σήμαινε ότι είχαμε πετύχει τον στόχο μας. Ένας από τους δημοσιογράφους παρατήρησε έναν συμμετέχοντα στο κυβερνητικό πρόγραμμα γονιδιώματος να κουνάει το κεφάλι του με θλίψη: «Φαίνεται ότι αυτοί οι απατεώνες θα κάνουν πραγματικά τα πάντα.» 1 Φύγαμε από το συνέδριο με μια νέα φόρτιση ενέργειας.
Έμειναν δύο σημαντικά προβλήματα προς επίλυση, τα οποία και τα δύο ήταν γνωστά σε εμάς. Ο πρώτος είναι ο τρόπος δημοσίευσης των αποτελεσμάτων. Παρά το μνημόνιο κατανόησης που είχαμε υπογράψει με τον Jerry Rubin, η επιχειρηματική μας ομάδα δεν αισθάνθηκε άνετα με την ιδέα της μεταφοράς πολύτιμων αποτελεσμάτων αλληλουχίας Drosophila στη GenBank. Πρότειναν να τοποθετηθούν τα αποτελέσματα της αλληλουχίας της μύγας φρούτων σε μια ξεχωριστή βάση δεδομένων στο Εθνικό Κέντρο Πληροφοριών Βιοτεχνολογίας, όπου ο καθένας θα μπορούσε να τα χρησιμοποιήσει υπό έναν όρο - όχι για εμπορικούς σκοπούς. Ο καυτερός, αλυσοκαπνιστής Michael Ashburner του Ευρωπαϊκού Ινστιτούτου Βιοπληροφορικής ήταν εξαιρετικά δυσαρεστημένος με αυτό. Πίστευε ότι η Celera είχε «εξαπατήσει τους πάντες» 2. (Έγραψε στον Ρούμπιν: «Τι διάολο συμβαίνει στο Σέλρα;» 3) Ο Κόλινς ήταν επίσης δυστυχισμένος, αλλά το πιο σημαντικό, το ίδιο και ο Τζέρι Ρούμπιν. Στο τέλος, έστειλα ακόμα τα αποτελέσματά μας στη GenBank.
Το δεύτερο πρόβλημα αφορούσε τη Drosophila - είχαμε τα αποτελέσματα της αλληλουχίας του γονιδιώματός της, αλλά δεν καταλάβαμε καθόλου τι σήμαιναν. Έπρεπε να τα αναλύσουμε αν θέλαμε να γράψουμε μια εργασία, όπως κάναμε πριν από τέσσερα χρόνια με τον Αιμοφίλο. Η ανάλυση και ο χαρακτηρισμός του γονιδιώματος της μύγας θα μπορούσε να διαρκέσει περισσότερο από ένα χρόνο - και δεν είχα αυτό το χρόνο, γιατί τώρα έπρεπε να εστιάσω στο ανθρώπινο γονιδίωμα. Αφού το συζητήσαμε με τον Jerry και τον Mark, αποφασίσαμε να εμπλέξουμε την επιστημονική κοινότητα στην εργασία για τη Drosophila, μετατρέποντάς το σε ένα συναρπαστικό επιστημονικό πρόβλημα, και έτσι να προχωρήσουμε γρήγορα το θέμα, κάνοντας μια διασκεδαστική διακοπές από τη βαρετή διαδικασία της περιγραφής του γονιδιώματος - σαν διεθνές τζάμπορι σκάουτινγκ. Το ονομάσαμε Genomic Jamboree και προσκαλέσαμε κορυφαίους επιστήμονες από όλο τον κόσμο να έρθουν στο Rockville για περίπου μια εβδομάδα ή δέκα ημέρες για να αναλύσουν το γονιδίωμα της μύγας. Με βάση τα αποτελέσματα που προέκυψαν, προγραμματίσαμε να γράψουμε μια σειρά άρθρων.
Η ιδέα άρεσε σε όλους. Ο Τζέρι άρχισε να στέλνει προσκλήσεις για την εκδήλωσή μας σε ομάδες κορυφαίων ερευνητών και οι ειδικοί της βιοπληροφορικής της Celera αποφάσισαν ποιοι υπολογιστές και προγράμματα θα χρειάζονταν για να κάνουν τη δουλειά των επιστημόνων όσο το δυνατόν πιο αποτελεσματική. Συμφωνήσαμε ότι η Celera θα πληρώσει τα έξοδα ταξιδιού και διαμονής τους. Μεταξύ των προσκεκλημένων ήταν οι πιο σκληροί επικριτές μου, αλλά ελπίζαμε ότι οι πολιτικές τους φιλοδοξίες δεν θα επηρέαζαν την επιτυχία του εγχειρήματός μας.
Τον Νοέμβριο, περίπου 40 ειδικοί της Drosophila έφτασαν σε εμάς, και ακόμη και για τους εχθρούς μας η προσφορά ήταν πολύ ελκυστική για να την αρνηθούν. Στην αρχή, όταν οι συμμετέχοντες συνειδητοποίησαν ότι έπρεπε να αναλύσουν περισσότερα από εκατό εκατομμύρια ζεύγη βάσεων γενετικού κώδικα μέσα σε λίγες μέρες, η κατάσταση ήταν αρκετά τεταμένη. Ενώ οι νεοαφιχθέντες επιστήμονες κοιμόντουσαν, το προσωπικό μου εργαζόταν όλο το εικοσιτετράωρο, αναπτύσσοντας προγράμματα για την επίλυση απρόβλεπτων προβλημάτων. Μέχρι το τέλος της τρίτης ημέρας, όταν αποδείχθηκε ότι τα νέα εργαλεία λογισμικού επιτρέπουν στους επιστήμονες, όπως είπε ένας από τους καλεσμένους μας, «να κάνουν εκπληκτικές ανακαλύψεις σε λίγες ώρες που προηγουμένως χρειάζονταν σχεδόν μια ζωή», η κατάσταση ηρέμησε. Κάθε μέρα στη μέση της ημέρας, με το σήμα του κινεζικού γκονγκ, μαζεύονταν όλοι μαζί για να συζητήσουν τα τελευταία αποτελέσματα, να λύσουν τρέχοντα προβλήματα και να καταρτίσουν ένα σχέδιο εργασίας για τον επόμενο γύρο.
Κάθε μέρα οι συζητήσεις γίνονταν όλο και πιο ενδιαφέρουσες. Χάρη στο Celera, οι καλεσμένοι μας είχαν την ευκαιρία να κοιτάξουν πρώτοι σε έναν νέο κόσμο και αυτό που αποκαλύφθηκε ξεπέρασε τις προσδοκίες. Σύντομα αποδείχθηκε ότι δεν είχαμε αρκετό χρόνο για να συζητήσουμε όλα όσα θέλαμε και να καταλάβουμε τι σήμαιναν όλα αυτά. Ο Μαρκ παρέταξε ένα εορταστικό δείπνο, το οποίο δεν κράτησε πολύ καθώς όλοι έσπευσαν γρήγορα πίσω στα εργαστήρια. Σύντομα τα μεσημεριανά γεύματα και τα δείπνα καταναλώθηκαν ακριβώς μπροστά από τις οθόνες των υπολογιστών με δεδομένα σχετικά με το γονιδίωμα της Drosophila να εμφανίζονται σε αυτές. Για πρώτη φορά, ανακαλύφθηκαν πολυαναμενόμενες οικογένειες γονιδίων υποδοχέων, μαζί με έναν εκπληκτικό αριθμό γονιδίων μύγας φρούτων παρόμοια με γονίδια ανθρώπινων ασθενειών. Κάθε ανακάλυψη συνοδευόταν από χαρούμενες κραυγές, σφυρίγματα και φιλικά χτυπήματα στον ώμο. Παραδόξως, εν μέσω της επιστημονικής μας γιορτής, ένα ζευγάρι βρήκε χρόνο να αρραβωνιαστεί.
Υπήρχε, ωστόσο, κάποια ανησυχία: κατά τη διάρκεια της εργασίας, οι επιστήμονες ανακάλυψαν μόνο περίπου 13 χιλιάδες γονίδια αντί για τα αναμενόμενα 20 χιλιάδες. Δεδομένου ότι το «χαμηλό» σκουλήκι C. elegans έχει περίπου 20 χιλιάδες γονίδια, πολλοί πίστευαν ότι η μύγα πρέπει να έχει περισσότερα από αυτά, αφού έχει 10 φορές περισσότερα κύτταρα και μάλιστα έχει νευρικό σύστημα. Υπήρχε ένας απλός τρόπος για να βεβαιωθούμε ότι δεν υπήρχε λάθος στους υπολογισμούς: πάρτε τα 2.500 γνωστά γονίδια της μύγας και δείτε πόσα από αυτά θα μπορούσαμε να βρούμε στην αλληλουχία μας. Μετά από προσεκτική ανάλυση, ο Michael Cherry από το Πανεπιστήμιο του Στάνφορντ ανέφερε ότι είχε βρει όλα τα γονίδια εκτός από έξι. Μετά από συζήτηση, αυτά τα έξι γονίδια ταξινομήθηκαν ως τεχνουργήματα. Το γεγονός ότι τα γονίδια εντοπίστηκαν χωρίς σφάλματα μας ενέπνευσε και μας έδωσε αυτοπεποίθηση. Μια κοινότητα χιλιάδων επιστημόνων αφοσιωμένων στην έρευνα της Drosophila είχε περάσει δεκαετίες παρακολουθώντας αυτά τα 2.500 γονίδια και τώρα 13.600 ήταν μπροστά τους στην οθόνη του υπολογιστή.
Κατά τη διάρκεια της αναπόφευκτης φωτογράφισης στο τέλος της δουλειάς, ήρθε μια αξέχαστη στιγμή: μετά το παραδοσιακό χτύπημα στον ώμο και φιλικές χειραψίες, ο Mike Ashburner έπεσε στα τέσσερα για να μπορέσω να απαθανατιστώ στη φωτογραφία με το πόδι μου στην πλάτη του. . Ήθελε λοιπόν -παρ' όλες τις αμφιβολίες και τον σκεπτικισμό του- να αποδώσει τα εύσημα στα επιτεύγματά μας. Ένας διάσημος γενετιστής και ερευνητής της Drosophila, βρήκε μάλιστα μια κατάλληλη λεζάντα για τη φωτογραφία: «Στακμένος στους ώμους ενός γίγαντα». (Είχε μια μάλλον αδύναμη φιγούρα.) «Ας δώσουμε τα εύσημα σε αυτούς που το αξίζουν», έγραψε αργότερα 4 . Οι αντίπαλοί μας προσπάθησαν να παρουσιάσουν τις καθυστερήσεις στη μεταφορά των αποτελεσμάτων της αλληλουχίας σε μια δημόσια βάση δεδομένων ως παρέκκλιση από τις υποσχέσεις μας, αλλά και αυτοί αναγκάστηκαν να παραδεχτούν ότι η συνάντηση είχε «μια εξαιρετικά πολύτιμη συμβολή στην παγκόσμια έρευνα για τις μύγες των φρούτων» 5 . Έχοντας βιώσει τι είναι η αληθινή «επιστημονική νιρβάνα», όλοι χώρισαν ως φίλοι.
Αποφασίσαμε να δημοσιεύσουμε τρεις μεγάλες εργασίες: μια για την αλληλουχία ολόκληρου του γονιδιώματος με τον Mike ως πρώτο συγγραφέα, μια για τη συναρμολόγηση γονιδιώματος με τον Gene ως πρώτο συγγραφέα και μια τρίτη για τη συγκριτική γονιδιωματική του σκουληκιού, της ζύμης και του ανθρώπινου γονιδιώματος με τον Jerry ως πρώτο συγγραφέας. Οι εργασίες υποβλήθηκαν στο Science τον Φεβρουάριο του 2000 και δημοσιεύτηκαν σε ειδικό τεύχος στις 24 Μαρτίου 2000, λιγότερο από ένα χρόνο μετά τη συνομιλία μου με τον Τζέρι Ρούμπιν στο Cold Spring Harbor. 6 Πριν από τη δημοσίευση, ο Τζέρι κανόνισε να μιλήσω στο ετήσιο Ερευνητικό Συνέδριο της Δροσόφιλας στο Πίτσμπουργκ, το οποίο παρακολούθησαν εκατοντάδες από τα πιο επιφανή άτομα στον τομέα. Σε κάθε καρέκλα στην αίθουσα, το προσωπικό μου τοποθέτησε ένα CD που περιείχε ολόκληρο το γονιδίωμα της Drosophila, καθώς και ανατυπώσεις των εργασιών μας που δημοσιεύτηκαν στο Science. Ο Τζέρι με σύστησε πολύ θερμά, διαβεβαιώνοντας το κοινό ότι είχα εκπληρώσει όλες τις υποχρεώσεις μου και ότι είχαμε συνεργαστεί καλά. Η ομιλία μου τελείωσε με μια αναφορά σε ορισμένες από τις έρευνες που έγιναν κατά τη διάρκεια της συνάντησης και έναν σύντομο σχολιασμό των δεδομένων στο CD. Το χειροκρότημα μετά την ομιλία μου ήταν τόσο εκπληκτικό και ευχάριστο όσο πριν από πέντε χρόνια όταν ο Χαμ και εγώ παρουσιάσαμε για πρώτη φορά το γονιδίωμα του Αιμοφίλου σε ένα συνέδριο μικροβιολογίας. Στη συνέχεια, οι εργασίες για το γονιδίωμα της Drosophila έγιναν οι πιο συχνά αναφερόμενες εργασίες στην ιστορία της επιστήμης.
Αν και χιλιάδες ερευνητές μύγας φρούτων σε όλο τον κόσμο ήταν ενθουσιασμένοι με τα αποτελέσματα, οι επικριτές μου γρήγορα πήγαν στην επίθεση. Ο John Sulston χαρακτήρισε την προσπάθεια αλληλουχίας του γονιδιώματος της μύγας αποτυχημένη, παρόλο που η αλληλουχία που λάβαμε ήταν πιο ολοκληρωμένη και ακριβέστερη από το αποτέλεσμα της επίπονης δεκαετούς προσπάθειάς του να καθορίσει την αλληλουχία του γονιδιώματος του σκουληκιού, η οποία χρειάστηκε άλλα τέσσερα χρόνια για να ολοκληρωθεί μετά τη δημοσίευση του σχεδίου στο Science. Ο συνάδελφος του Sulston, Maynard Olson, χαρακτήρισε την αλληλουχία του γονιδιώματος της Drosophila «ντροπή» που θα έπρεπε να διευθετήσει το κυβερνητικό πρόγραμμα ανθρώπινου γονιδιώματος, «με τη χάρη» του Celera. Στην πραγματικότητα, η ομάδα του Jerry Rubin μπόρεσε να κλείσει γρήγορα τα εναπομείναντα κενά στην αλληλουχία δημοσιεύοντας και αναλύοντας συγκριτικά το ήδη αλληλουχημένο γονιδίωμα σε λιγότερο από δύο χρόνια. Αυτά τα δεδομένα επιβεβαίωσαν ότι είχαμε 1–2 σφάλματα ανά 10 kb σε ολόκληρο το γονιδίωμα και λιγότερο από 1 σφάλμα ανά 50 kb στο λειτουργικό (ευχρωματικό) γονιδίωμα.
Ωστόσο, παρά τη γενική αναγνώριση του έργου Drosophila, οι εντάσεις στη σχέση μου με τον Tony White έφτασαν σε πυρετό το καλοκαίρι του 1999. Ο White δεν μπορούσε να συμβιβαστεί με την προσοχή που έδινε ο Τύπος στο άτομό μου. Κάθε φορά που ερχόταν στο Celera, περνούσε από αντίγραφα άρθρων για τα επιτεύγματά μας κρεμασμένα στους τοίχους στο διάδρομο δίπλα στο γραφείο μου. Και εδώ μεγεθύναμε ένα από αυτά - το εξώφυλλο του κυριακάτικου συμπληρώματος της εφημερίδας USA Today. Σε αυτό, υπό τον τίτλο «Θα κάνει αυτός ο τυχοδιώκτης τη μεγαλύτερη επιστημονική ανακάλυψη της εποχής μας;» Το 7 με έδειξε, με ένα μπλε καρό πουκάμισο, να σταυρώνω τα πόδια μου, και γύρω μου ο Κοπέρνικος, ο Γαλιλαίος, ο Νεύτωνας και ο Αϊνστάιν επέπλεαν στον αέρα - και κανένα σημάδι από τον Λευκό.
Κάθε μέρα, ο γραμματέας Τύπου του τηλεφωνούσε για να δει αν ο Τόνι μπορούσε να λάβει μέρος στη φαινομενικά ατελείωτη ροή συνεντεύξεων που λάμβανε χώρα στο Celera. Ηρέμησε λίγο - και μάλιστα μόνο για λίγο, όταν τον επόμενο χρόνο κατάφερε να τοποθετήσει τη φωτογραφία του στο εξώφυλλο του περιοδικού Forbes ως ο άνθρωπος που μπόρεσε να αυξήσει την κεφαλαιοποίηση του PerkinElmer από 1,5 δισεκατομμύρια δολάρια σε 24 δισεκατομμύρια δολάρια 8 . («Ο Τόνι Γουάιτ μετέτρεψε τον φτωχό Πέρκιν Έλμερ σε σύλληψη γονιδίων υψηλής τεχνολογίας.») Ο Τόνι στοιχειωνόταν επίσης από τις κοινωνικές μου δραστηριότητες.
Έκανα μια ομιλία μια φορά την εβδομάδα, δεχόμενος ένα μικρό κλάσμα από τον τεράστιο αριθμό των προσκλήσεων που λάμβανα συνεχώς επειδή ο κόσμος ήθελε να μάθει για τη δουλειά μας. Ο Tony παραπονέθηκε ακόμη και στο διοικητικό συμβούλιο της PerkinElmer, που τότε μετονομάστηκε σε PE Corporation, ότι τα ταξίδια και οι εμφανίσεις μου παραβίαζαν τους εταιρικούς κανόνες. Κατά τη διάρκεια δύο εβδομάδων διακοπών (με δικά μου έξοδα) στο σπίτι μου στο Cape Cod, ο Tony πέταξε στο Celera με τον CFO Dennis Winger και τον γενικό σύμβουλο της Applera William Sauch για να πάρει συνέντευξη από τους κορυφαίους υπαλλήλους μου σχετικά με την «αποτελεσματικότητα διαχείρισης του Venter». Ήλπιζαν να μαζέψουν αρκετό χώμα για να δικαιολογήσουν την απόλυσή μου. Ο Γουάιτ σοκαρίστηκε όταν όλοι είπαν ότι αν τα παράτησα, θα τα παράτησαν κι εκείνοι. Αυτό προκάλεσε μεγάλη ένταση στην ομάδα μας, αλλά μας έφερε και πιο κοντά από ποτέ. Ήμασταν έτοιμοι να πανηγυρίσουμε κάθε νίκη σαν να ήταν η τελευταία μας.
Μετά τη δημοσίευση της αλληλουχίας του γονιδιώματος της μύγας - μέχρι τότε της μεγαλύτερης αλληλουχίας στην ιστορία - ο Τζιν, ο Χαμ, ο Μαρκ και εγώ είπαμε ότι σταθήκαμε στον Τόνι Γουάιτ αρκετό καιρό ώστε να αναγνωριστεί η επιτυχία μας. Έχουμε αποδείξει ότι η μέθοδός μας θα λειτουργήσει και κατά τον προσδιορισμό της αλληλουχίας του ανθρώπινου γονιδιώματος. Ακόμα κι αν ο Tony White σταματούσε να χρηματοδοτεί την επόμενη μέρα, ξέραμε ότι το κύριο επίτευγμά μας θα παρέμενε μαζί μας. Περισσότερο από οτιδήποτε άλλο, ήθελα να φύγω από την Celera και να μην έχω να ασχοληθώ με τον Tony White, αλλά επειδή ήθελα να ακολουθήσω το γονιδίωμα του Homo sapiens ακόμα περισσότερο, έπρεπε να κάνω έναν συμβιβασμό. Προσπάθησα όσο καλύτερα μπορούσα να ευχαριστήσω τον White, απλώς για να συνεχίσω τη δουλειά και να ολοκληρώσω το σχέδιό μου.
Σημειώσεις
1. Shreeve J. The Genome War: How Craig Venter Tried to Capture the Code of Life and Save the World (Νέα Υόρκη: Ballantine, 2005), σελ. 285.
2. Ashburner M. Won for All: How the Drosophila Genome Was Sequenced (Cold Spring Harbor Laboratory Press, 2006), σελ. 45.
3. Shreeve J. The Genome War, σελ. 300.
4. Ashburner M. Won for All, σελ. 55.
5. Sulston J., Ferry G. The Common Thread (Λονδίνο: Corgi, 2003), σελ. 232.
6. Adams M. D., Celniker S. E. et al. "The Genome Sequence of Drosophila Melanogaster", Science, αρ. 287, 2185–95, 24 Μαρτίου 2000.
7. Gillis J. «Θα ξεκλειδώσει αυτό το MAVERICK τη μεγαλύτερη επιστημονική ανακάλυψη της εποχής του; Κοπέρνικος, Νεύτωνας, Αϊνστάιν και ΒΕΝΤΕΡ;», Σαββατοκύριακο ΗΠΑ, 29–31 Ιανουαρίου 1999.
8. Ross P. E. “Gene Machine”, Forbes, 21 Φεβρουαρίου 2000.
Κρεγκ Βέντερ