Schemat pozyskiwania hybryd pierwszej generacji. Pierwszym prawem Mendla jest prawo dominacji. Prawo czystości gamet i jego cytologiczne uzasadnienie
6. Czym jest ontogeneza. Jakie są okresy ontogenezy? Podaj krótki opis rozwoju postembrionalnego.
1. Poziomy organizacji dzikich zwierząt (lista).
2. Wymień organelle komórkowe dwubłonowe i bezbłonowe. Wskaż cechy ich struktury i funkcji.
3. Jakie są zalety procesu rozmnażania płciowego nad bezpłciowym.
4. Trzecie prawo Mendla. Wskaż proporcję potomstwa według genotypu i fenotypu.
5. Wymień ośrodki pochodzenia i różnorodności roślin uprawnych wg Wawilowa. Gdzie przechowywana jest kolekcja nasion roślin rosnących na Ziemi, jakie jest jej znaczenie.
6. Czym jest ontogeneza. Jakie są okresy ontogenezy? Podaj krótki opis rozwoju postembrionalnego.
ciało gospodarza. Środki zapobiegawcze.
2. Siły napędowe ewolucji i ich związek.
5. Określ powiększenie mikroskopu. Przygotuj go do pracy.
Numer biletu 6
1. Układ krążenia kręgowców.
2.1. Skrzyżowanie monohybrydowe.
2. Pierwsze i drugie prawo Mendla.
6. Korzystając z materiału zielnikowego, pogrupuj rośliny w rodziny.
Numer biletu 7
1. Oddziały układu nerwowego: centralny i obwodowy. 2. Siedlisko organizmów.
Biologia, pomoc !!!1. Nauka badająca wzorce przekazywania cech przez dziedziczenie
2. Najpierw sformułowano wzorce genetyczne
3. Sparowane chromosomy, z których każdy trafia do ciała od jednego z rodziców
4. Chromosomy zawierające zestaw identycznych genów
5. Chromosomy zlokalizowane w tym samym organizmie, ale różniące się wielkością, kształtem i zestawem genów
6. Geny zlokalizowane w tych samych regionach chromosomów homologicznych, odpowiedzialne za syntezę tych samych białek, ale o innej sekwencji nukleotydowej
7. Jaka będzie nazwa genów kodujących to samo białko (na przykład hemoglobina lub melanina), ale zawierających inną sekwencję aminokwasową
8. W przypadku krzyżowania monohybrydowego znaki są badane
9. Zgodnie z genem koloru (w eksperymentach Mendla) groch posiadał szereg cech
10. Zygota, która otrzymała dwa różne allele
11. Zygota, która otrzymała oba te same allele
12. Gen oznaczony po przekreśleniu małą literą
13. Jak nazywa się allel, oznaczany przy skrzyżowaniu dużej (dużej) litery?
14. Allel, który fenotypowo pojawia się tylko w stanie homozygotycznym
15. Allel, który w każdym przypadku objawia się zewnętrznie, jeśli w ogóle jest obecny w genotypie osobnika
16. Genotyp zygoty oznaczony AA
17. Genotyp zygoty oznaczony Aa
18. Genotyp zygoty, oznaczony aa
19. Zestaw zewnętrznych znaków ciała
20. Całość wszystkich dziedzicznych cech organizmu
21. Zbiór wszystkich genów gatunku lub populacji
22. Podział mieszańców w drugim pokoleniu przebiega według wzoru 3:1
23. Genotyp form rodzicielskich grochu („czyste linie”) w eksperymentach Mendla
24. Genotyp potomków (hybrydy I pokolenia) grochu w eksperymentach Mendla
25. Kolor grochu pierwszej generacji
26. F1 Mendel uzyskany przez zapylenie
27. F2 Mendel uzyskany przez zapylanie
28. Rodzaj cięcia przy krzyżowaniu heterozygota i homozygota na recesywny
29. Formy monoheterozygoty ... odmiany gamet
30. Hybrydy z krzyżowaniem monohybrydowym i formą pełnej dominacji ... klasy fenotypowe
31. Prawa Mendla dotyczyły tego rodzaju dziedziczenia
32. Geny niealleliczne zlokalizowane na tym samym chromosomie, jeśli nie dochodzi do krzyżowania, są dziedziczone
33. Autor chromosomowej teorii dziedziczności
34. Chromosomy różniące się wielkością i kształtem u mężczyzn i kobiet
35. Chromosomy, które nie różnią się wielkością i kształtem
36. Znajdują się geny „związane z płcią”
37. Przykładami są hemofilia i ślepota barw u ludzi oraz dziedziczenie koloru sierści u kotów
38. Znajdują się geny tych cech
39. Samce ssaków i Drosophila są heterozygotyczne pod względem cech związanych z płcią
40. Przypadkowe zmiany w materiale genetycznym osobnika
41. Zmiana liczby chromosomów
42. Zmiana jednego lub więcej nukleotydów
43. Zmiany w sekwencji aminokwasowej białek
44. Autor prawa szeregu homologicznego zmienności dziedzicznej
45. Główna metoda genetyki, niemożliwa do zastosowania w genetyce człowieka
46. Metoda genetyki człowieka, oparta na barwieniu chromosomów i późniejszym badaniu ich wielkości i kształtu
47. Metoda genetyki człowieka, oparta na analizie rodowodów i śledzeniu przenoszenia określonej cechy
48. Metoda genetyki człowieka oparta na analizie przejawów fenotypowych cech u bliźniąt jednojajowych
49. Metoda genetyki człowieka, oparta na analizie aktywności enzymatycznej białek katalizujących ważne procesy fizjologiczne
50. Substancje powodujące mutacje
Wstęp.
Genetyka to nauka badająca prawa dziedziczności i zmienności organizmów żywych.
Człowiek od dawna zwraca uwagę na trzy zjawiska związane z dziedzicznością: po pierwsze, podobieństwo cech potomków i rodziców; po drugie, różnice między niektórymi (czasami wieloma) cechami potomstwa od odpowiadających im cech rodzicielskich; po trzecie, pojawienie się u potomstwa postaci, które były tylko u odległych przodków. Ciągłość cech międzypokoleniowych zapewnia proces zapłodnienia. Od niepamiętnych czasów człowiek spontanicznie wykorzystywał właściwości dziedziczności do celów praktycznych - do hodowli odmian roślin uprawnych i ras zwierząt domowych.
Pierwsze idee dotyczące mechanizmu dziedziczenia wyrazili starożytni greccy naukowcy Demokryt, Hipokrates, Platon, Arystoteles. Autor pierwszej naukowej teorii ewolucji J.-B. Lamarck wykorzystał idee starożytnych greckich naukowców, aby wyjaśnić to, co postulował na przełomie XVIII i XIX wieku. zasada przekazywania potomstwu nowych cech nabytych w życiu jednostki. C. Darwin wysunął teorię pangenezy, wyjaśniając dziedziczenie cech nabytych
Definicja Karola Darwina dziedziczność jako właściwość wszystkich żywych organizmów do przekazywania ich cech i właściwości z pokolenia na pokolenie, oraz zmienność jako właściwość wszystkich żywych organizmów do nabywania nowych cech w procesie indywidualnego rozwoju.
Cechy są dziedziczone poprzez reprodukcję. Wraz z rozmnażaniem płciowym w wyniku zapłodnienia powstają nowe pokolenia. Materialne podstawy dziedziczności zawarte są w komórkach zarodkowych. W przypadku rozmnażania bezpłciowego lub wegetatywnego nowe pokolenie rozwija się albo z jednokomórkowych zarodników, albo z formacji wielokomórkowych. A przy tych formach rozmnażania połączenie między pokoleniami odbywa się za pośrednictwem komórek, które zawierają materialne podstawy dziedziczności (elementy elementarne dziedziczności) - geny - są odcinkami DNA chromosomów.
Zestaw genów, które organizm otrzymuje od rodziców, stanowi jego genotyp. Całość cech zewnętrznych i wewnętrznych jest fenotypem. Fenotyp rozwija się w wyniku interakcji genotypu i warunków środowiskowych. Tak czy inaczej, podstawą są cechy niosące geny.
Wzorce, według których znaki są przekazywane z pokolenia na pokolenie, zostały po raz pierwszy odkryte przez wielkiego czeskiego naukowca Gregora Mendla. Odkrył i sformułował trzy prawa dziedziczenia, które stanowiły podstawę współczesnej genetyki.
Życie i badania naukowe Gregora Johanna Mendla.
Morawski mnich i genetyk roślin. Johann Mendel urodził się w 1822 r. w miejscowości Heinzendorf (obecnie Gincice w Czechach), gdzie jego ojciec był właścicielem małej chłopskiej działki. Gregor Mendel, według zeznań tych, którzy go znali, był rzeczywiście miłym i miłym człowiekiem. Po ukończeniu szkoły podstawowej w tutejszej wiejskiej szkole, a później, po ukończeniu Kolegium Pijarów w Leipniku, został przyjęty w 1834 r. do I klasy gimnazjum Cesarsko-Królewskiego w Troppown. Cztery lata później rodzice Johanna, w wyniku zbiegu wielu nieszczęśliwych wydarzeń, które szybko następowały po sobie, zostali całkowicie pozbawieni możliwości zwrotu niezbędnych wydatków związanych z edukacją, a ich syn, mając wówczas zaledwie 16 lat, został zmuszony całkowicie samodzielnie zająć się własnym utrzymaniem.... W 1843 roku Mendel został przyjęty do klasztoru augustianów św. Tomasza w Altbrunn, gdzie przyjął imię Gregor. W 1846 r. Mendel uczęszczał również na wykłady o rolnictwie, ogrodnictwie i uprawie winorośli w Instytucie Filozoficznym w Brunn. W 1848 r. po ukończeniu kursu teologicznego Mendel otrzymał z głębokim szacunkiem pozwolenie na studia do egzaminów doktorskich. Gdy w następnym roku umocnił chęć przesłuchania, otrzymał rozkaz zajęcia miejsca zwolennika cesarsko-królewskiego gimnazjum w Znaim, za czym z radością podążał.
W 1851 r. opat klasztoru wysłał Mendla na studia na uniwersytecie wiedeńskim, gdzie studiował m.in. botanikę. Po ukończeniu studiów Mendel nauczał przedmiotów ścisłych w miejscowej szkole. Dzięki temu krokowi jego sytuacja finansowa radykalnie się zmieniła. W dobroczynnym dobrobycie fizycznej egzystencji, tak niezbędnej w każdym dążeniu, odwaga i siła powróciły do niego z głębokim szacunkiem, a przez cały rok próbny studiował przepisane klasyczne przedmioty z wielką pilnością i miłością. W wolnych chwilach zajmował się niewielką kolekcją botaniczną i mineralogiczną, którą miał do dyspozycji w klasztorze. Jego pasja do dziedziny nauk przyrodniczych stawała się tym większa, im więcej miał możliwości, by mu się poddać. Chociaż osoba wspomniana na tych lekcjach została pozbawiona jakiegokolwiek przewodnictwa, a droga samouczenia tutaj, jak w żadnej innej nauce, jest trudna i powoli prowadzi do celu, to jednak w tym czasie Mendel nabrał takiego zamiłowania do badania przyrody, że nie szczędził wysiłków, aby wypełnić zmienione luki poprzez samodzielną naukę i stosowanie się do rad ludzi z praktycznym doświadczeniem. 3 kwietnia 1851 r. „korpus nauczycielski” szkoły postanowił zaprosić pana Gregora Mendla, kanonika klasztoru św. Sukcesy pomologiczne Gregora Mendla przyniosły mu prawo do tytułu gwiazdy i tymczasowego stanowiska zwolennika historii naturalnej w klasie przygotowawczej Technikum. W pierwszym semestrze studiów studiował tylko dziesięć godzin tygodniowo i tylko z Dopplerem. W drugim semestrze uczył się dwadzieścia godzin tygodniowo. Spośród nich dziesięć - z fizyki z Dopplerem, pięć tygodniowo - z zoologii z Rudolfem Knerem. Jedenaście godzin tygodniowo - botanika z prof. Fenzlem: oprócz wykładów z morfologii i taksonomii przeszedł także specjalne warsztaty z opisu i definicji roślin. W trzecim semestrze zapisał się na 32 godziny zajęć tygodniowo: dziesięć godzin z fizyki w Dopplerze, dziesięć z chemii w Rottenbacher: chemia ogólna, chemia medyczna, chemia farmakologiczna i warsztaty z chemii analitycznej. Pięć dla zoologii Knera. Sześć godzin treningu z Ungerem, jednym z pierwszych cytologów na świecie. W swoich laboratoriach studiował anatomię i fizjologię roślin oraz wziął udział w warsztatach z technik mikroskopowych. I znowu - raz w tygodniu na Wydziale Matematyki - warsztaty z logarytmu i trygonometrii.
1850, życie szło dobrze. Mendel mógł już sam się utrzymać i cieszył się wielkim szacunkiem kolegów, bo dobrze radził sobie ze swoimi obowiązkami i bardzo przyjemnie się z nim rozmawiało. Jego uczniowie kochali go.
W 1851 roku Gregor Mendel porzucił kardynalną kwestię biologii – problem zmienności i dziedziczności. Wtedy to zaczął przeprowadzać eksperymenty z ukierunkowaną uprawą roślin. Mendel przywiózł różne rośliny z dalekich i bliskich okolic Brunn. Uprawiał rośliny w grupach w specjalnie dla każdego z nich wydzielonej części ogrodu klasztornego w różnych warunkach zewnętrznych. Zajmował się żmudnymi obserwacjami meteorologicznymi. Większość swoich doświadczeń i obserwacji Gregor prowadził z groszkiem, który od 1854 r. co wiosnę siał każdej wiosny w małym ogródku pod oknami prałatury. Okazało się, że łatwo przeprowadzić klarowny eksperyment hybrydyzacji na grochu. Aby to zrobić, wystarczy otworzyć pęsetą duży, choć jeszcze niedojrzały kwiat, odciąć pylniki i samodzielnie określić „parę” do skrzyżowania. Ponieważ samozapylenie jest wykluczone, odmiany grochu są z reguły „czystymi liniami” o stałych cechach, które nie zmieniają się z pokolenia na pokolenie, co jest bardzo wyraźnie zarysowane. Mendel wyróżnił cechy, które determinowały różnice międzyodmianowe: kolor skórki ziaren dojrzałych i osobno ziaren niedojrzałych, kształt grochu dojrzałego, kolor „białka” (bielmo), długość osi łodygi , położenie i kolor pąków. W eksperymencie wykorzystał ponad trzydzieści odmian, a każda z odmian została wcześniej poddana dwuletniemu testowi na „stałość”, na „stałość cech”, na „czystość krwi” – w 1854 i 1855 roku. Eksperymenty z groszkiem trwały osiem lat. Setki razy w ośmiu kwitnieniach własnymi rękami ostrożnie odcinał pylniki i, wpisując pyłek z pręcików kwiatu innej odmiany na pęsetach, nakładał go na piętno słupka. Dziesięć tysięcy paszportów wydano dla dziesięciu tysięcy roślin uzyskanych w wyniku krzyżówek iz mieszańców samopylnych. Wpisy w nich są zgrabne: kiedy uprawiano roślinę rodzicielską, jakie miała kwiaty, czyje pyłek został nawożony, jaki groch - żółty czy zielony, gładki czy pomarszczony - uzyskano, które kwiaty - kolor na brzegach, kolor w centrum - rozkwitło, gdy otrzymano nasiona, ile z nich jest żółtych, ile jest zielonych, okrągłych, pomarszczonych, ile z nich wybiera się do sadzenia, kiedy są sadzone i tak dalej.
Efektem jego badań był raport „Eksperymenty na mieszańcach roślin”, który w 1865 r. przeczytał brunniański przyrodnik. Raport mówi: „Powodem rozpoczęcia eksperymentów, którym poświęcony jest ten artykuł, było sztuczne krzyżowanie roślin ozdobnych, które przeprowadzono w celu uzyskania nowych form, różniących się kolorem. Rozpoczęcie dalszych eksperymentów w celu prześledzenia rozwoju mieszańców u ich potomstwa dało impuls do rzucającej się w oczy regularności, z jaką formy mieszańcowe stale powracały do form przodków. Jak to często bywa w historii nauki, twórczość Mendla nie zyskała od razu należytego uznania u współczesnych. Wyniki jego eksperymentów zostały opublikowane na spotkaniu Towarzystwa Nauk Przyrodniczych miasta Brunn, a następnie opublikowane w czasopiśmie tego Towarzystwa, ale idee Mendla w tym czasie nie znalazły poparcia. Numer pisma opisującego rewolucyjne dzieło Mendla od trzydziestu lat kurzy się w bibliotekach. Dopiero pod koniec XIX wieku naukowcy zajmujący się problematyką dziedziczności odkryli dzieła Mendla i udało mu się uzyskać (już pośmiertnie) zasłużone uznanie.
Pierwsze prawo Mendla
Nazywa się krzyżowanie dwóch organizmów hybrydyzacja, potomstwo ze skrzyżowania dwóch osobników o różnej dziedziczności nazywa się hybrydowy, i osobna osoba - hybryda.Mononukleozahybrydowy nazywa się krzyżowaniem dwóch organizmów, które różnią się od siebie jedną parą alternatywnych (wzajemnie wykluczających się) cech. W związku z tym przy takim krzyżowaniu śledzone są wzorce dziedziczenia tylko dwóch cech, których rozwój wynika z pary genów allelicznych. Wszystkie inne znaki charakterystyczne dla tych organizmów nie są brane pod uwagę.
Jeśli krzyżujesz rośliny grochu z żółtymi i zielonymi nasionami, wszystkie powstałe hybrydy będą miały żółte nasiona. Ten sam obraz obserwuje się podczas krzyżowania roślin o gładkim i pomarszczonym kształcie nasion; całe potomstwo pierwszego pokolenia będzie miało gładki kształt nasion:
Dlatego nazwano pierwsze prawo Mendla Prawo jednorodności hybryd pierwszej generacji.
Jeśli użyjemy terminów, które pojawiły się wiele lat po pracy Mendla, to możemy powiedzieć, że komórki roślin grochu jednej odmiany zawierają dwa geny tylko koloru żółtego, a geny roślin innej odmiany - dwa geny tylko koloru zielonego. Geny odpowiedzialne za rozwój jednej cechy (na przykład koloru nasion) nazywane są geny alleliczne... W konsekwencji w hybrydzie pierwszego pokolenia rozwija się tylko jedna z każdej pary cech alternatywnych. Drugi objaw wydaje się zanikać, nie pojawia się. Zjawisko dominacji w hybrydzie zznak jednego z rodziców G. Mendla nazwany dominracjonowanie. Nazwano cechę, która przejawia się w hybrydzie pierwszego pokolenia i hamuje rozwój innej cechy dominujący, i odwrotnie, czyli stłumiony, znak - recesywny. Gregor Mendel postanowił zastąpić opis cech roślin abstrakcyjnym kodem „A, B, C, D, E, F, G” oraz „a, b, c, d, e, f, g”, a następnie Los jednej pary cech przeniósł się do oglądania dwóch, trzech, czterech par jednocześnie. Duże A, B, C, D, E, F, G zaznaczył cechy dominujące; małe a, b, c, d, e, f, g - recesywne. Jeśli w genotypie organizmu (zygoty) występują dwa identyczne geny alleliczne, oba są dominujące lub oba są recesywne (AA lub aa), taki organizm nazywa się homozygotyczny. Jeśli z pary genów allelicznych jeden jest dominujący, a drugi recesywny (Aa), wtedy taki organizm nazywa się heterozygotyczny.
Prawo podziału, czyli drugie prawo Mendla.
Jeżeli potomkowie pierwszego pokolenia, identyczni w badanej cesze, zostaną ze sobą skrzyżowani, to w drugim pokoleniu cechy obojga rodziców pojawiają się w określonej proporcji liczbowej: 3/4 osobników będzie miało cechę dominującą, ¼ recesywny:

Zjawisko, w którym krzyżowanie heterozygotycznychosobniki prowadzą do powstania potomstwa, z których częśćrogo nosi cechę dominującą, a część- recesywnyny, nazywa się dzieleniem. W konsekwencji cecha recesywna u mieszańców pierwszego pokolenia nie zniknęła, a jedynie została stłumiona i ujawni się w drugim pokoleniu mieszańców.
Hipoteza czystości gamet. Mendel zasugerował, że podczas formowania się mieszańców czynniki dziedziczne nie mieszają się, ale pozostają niezmienione. W hybrydzie występują oba czynniki – dominujący i recesywny, przy czym dominujący czynnik dziedziczny przejawia się w postaci cechy, natomiast czynnik recesywny jest stłumiony. Komunikacja między pokoleniami podczas rozmnażania płciowego odbywa się za pośrednictwem komórek płciowych - mammeta. Dlatego należy założyć, że każda gameta zawiera tylko jeden czynnik z pary. Następnie, podczas zapłodnienia, fuzja dwóch gamet, z których każda niesie recesywny czynnik dziedziczny, doprowadzi do powstania organizmu z cechą recesywną, która objawia się fenotypowo. Fuzja gamet, z których każda zawiera czynnik dominujący, lub dwie gamety, z których jedna zawiera czynnik dominujący, a druga czynnik recesywny, prowadzi do rozwoju organizmu z cechą dominującą.
Mendel tłumaczył podział potomstwa podczas krzyżowania osobników heterozygotycznych faktem, że gamety są czyste genetycznie, to znaczy niosą tylko jeden gen z pary allelicznej. Hipoteza(teraz nazywany prawem) czystośćgamety można sformułować w następujący sposób: podczas tworzenia komórek zarodkowych do każdej gamety dostaje się tylko jeden gen z pary alleli.
Dlaczego i jak to się dzieje? Wiadomo, że każda komórka ciała ma dokładnie taki sam diploidalny zestaw chromosomów. Dwa homologiczne chromosomy zawierają dwa identyczne geny. Genetycznie „czyste” gamety powstają w następujący sposób: gdy łączą się gamety męskie i żeńskie, uzyskuje się hybrydę z diploidalnym (podwójnym) zestawem chromosomów.
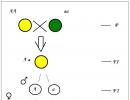
Jak widać na diagramie (Załącznik 2), zygota otrzymuje połowę chromosomów od organizmu ojcowskiego, a połowę od matki.
W procesie tworzenia gamet w hybrydzie chromosomy homologiczne podczas pierwszego podziału mejotycznego również wchodzą do różnych komórek.
W przypadku tej pary allelicznej powstają dwie odmiany gamet. Podczas zapłodnienia gamety niosące te same lub różne allele przypadkowo spotykają się ze sobą. Ze względu na prawdopodobieństwo statystyczne, przy odpowiednio dużej liczbie gamet u potomstwa, 25% genotypów będzie homozygotycznych dominujących, 50% - heterozygotycznych, 25% - homozygotycznych recesywnych, czyli stosunek 1AA: 2Aa: 1AA.
Odpowiednio, zgodnie z fenotypem, potomstwo drugiego pokolenia podczas krzyżowania monohybrydowego rozkłada się w proporcji 3:1 (¾ osobników z cechą dominującą, ¼ osobników z cechą recesywną).
Podstawą cytologiczną rozszczepiania cech w krzyżowaniu monohybrydowym jest dywergencja chromosomów homologicznych do różnych biegunów komórki i powstawanie haploidalnych komórek płciowych w mejozie.
W omówionych powyżej przykładach zasada jednorodności wyrażała się w tym, że wszystkie mieszańce były zewnętrznie podobne do jednego z rodziców. Nie zawsze tak jest. Często znaki w formach heterozygotycznych mają charakter pośredni, tj. dominacja może nie być pełna. Schemat skrzyżowania dwóch dziedzicznych form rośliny Night Beauty:


Jedna z nich ma czerwone kwiaty (i to jest cecha dominująca), a druga jest biała. Diagram pokazuje, że wszystkie hybrydy pierwszej generacji mają różowe kwiaty. W drugiej generacji rozszczepienie następuje w stosunku 1: 2: 1, tj. jeden czerwony kwiat (homozygota), dwa różowe kwiaty (heterozygota), jeden biały (homozygota). Zjawisko to nazywa się niepełną dominacją.
W przypadku niepełnej dominacji, dominujący gen w stanie heterozygotycznym nie zawsze całkowicie tłumi gen recesywny. W niektórych przypadkach hybryda fi nie odtwarza w pełni żadnej z cech rodzicielskich, a cecha ta ma charakter pośredni, z mniejszym lub większym odchyleniem w kierunku stanu dominującego lub recesywnego. Ale wszystkie osobniki tego pokolenia są jednolite w tej charakterystyce. Niepełna dominacja jest powszechna. Stwierdzono to w badaniach nad dziedziczeniem koloru kwiatów u lwiej paszczy, koloru wełny u bydła i owiec, cech biochemicznych u ludzi itp. Cechy pośrednie wynikające z niepełnej dominacji często reprezentują wartość estetyczną lub materialną dla człowieka. Powstaje pytanie: czy można rozmnażać się przez selekcję, na przykład odmianę nocnego piękna o różowym kolorze kwiatów? Oczywiście nie, ponieważ ta cecha rozwija się tylko u heterozygot i przy ich krzyżowaniu zawsze następuje rozszczepienie:
Niepełna dominacja jest powszechna. Stwierdzono to w badaniach nad dziedziczeniem koloru kwiatów u lwiej paszczy, koloru wełny u bydła i owiec, cech biochemicznych u ludzi itp. Cechy pośrednie wynikające z niepełnej dominacji często reprezentują wartość estetyczną lub materialną dla człowieka. Powstaje pytanie: czy można rozmnażać się przez selekcję, na przykład odmianę nocnego piękna o różowym kolorze kwiatów? Oczywiście, że nie, ponieważ ta cecha rozwija się tylko u heterozygot i przy ich krzyżowaniu zawsze następuje rozszczepienie.
Prawo niezależnej kombinacji, czyli trzeciePrawo Mendla.
Badanie Mendla dziedziczenia jednej pary alleli pozwoliło ustalić szereg ważnych wzorców genetycznych: zjawisko dominacji, niezmienność alleli recesywnych w mieszańcach, podział potomstwa mieszańców w stosunku 3:1, a także założyć, że gamety są genetycznie czyste, to znaczy zawierają tylko jeden gen z par allelicznych. Jednak organizmy różnią się wieloma genami. Możliwe jest ustalenie wzorców dziedziczenia dwóch par cech alternatywnych i więcej poprzez dwuhybrydowy lub krzyżowanie wielohybrydowe, tj. krzyżowanie form rodzicielskich różniących się dwiema parami cech.
Do krzyżowania dihybrydów Mendel wybrał homozygotyczne rośliny grochu, które różnią się dwoma wskaźnikami - kolorem nasion (żółty, zielony) i kształtem nasion (gładkie, pomarszczone). Cechy dominujące - żółte zabarwienie (A) i gładki kształt (V) posiew. Każda roślina tworzy jedną odmianę gamet dla badanych alleli: Kiedy gamety się łączą, całe potomstwo będzie jednolite:


|
Krata Pannet |
||||||

Łatwo obliczyć, że według fenotypu potomstwo dzieli się na 4 grupy: 9 żółtych gładkich, 3 żółtych pomarszczonych, 3 zielonych gładkich, 1 żółtej pomarszczonej (9:3:3:1). Jeśli weźmiemy pod uwagę wyniki rozszczepienia dla każdej pary cech z osobna, to okaże się, że stosunek liczby nasion żółtych do liczby nasion zielonych oraz stosunek nasion gładkich do pomarszczonych dla każdej pary wynosi 3:1 . Można to wyrazić algebraicznie jako kwadrat dwumianu
(3 + 1) ² = 3² + 2 · 3 + 1² lub 9 + 3 + 3 + 1
Zatem w krzyżówce dihybrydowej każda para cech podczas rozdzielenia u potomstwa zachowuje się tak samo jak w krzyżówce monohybrydowej, czyli niezależnie od drugiej pary cech.
Podczas zapłodnienia gamety łączy się według zasad losowych kombinacji, ale z równym prawdopodobieństwem dla każdej z nich. W powstałych zygotach powstają różne kombinacje genów. Teraz możemy sformułować trzecie prawo Mendla: podczas krzyżowania dwóch homozygotycznych osobników, różniących sięoddzieleni od siebie dwie lub więcej par alternatywnych cech, genów iodpowiadające im cechy są dziedziczone niezależnieod siebie i są połączone we wszystkie możliwekombinacje.
Prawa Mendla służą jako podstawa do analizy rozszczepienia w bardziej złożonych przypadkach: gdy osobniki różnią się trzema, czterema parami znaków lub więcej.
Warunki przestrzegania praw spadkowych Mendla
Prawa odkryte przez Gregora Mendla nie zawsze mają zastosowanie w genetyce. Istnieje wiele warunków przestrzegania praw Mendla. W takich przypadkach istnieją inne prawa (na przykład: prawo Morgana) lub wyjaśnienia.
Sformułujmy podstawowe warunki przestrzegania praw dziedziczenia.
Aby zachować zgodność z prawem jednorodności hybryd pierwszej generacji, konieczne jest, aby:
organizmy rodzicielskie były homozygotyczne;
geny różnych alleli znajdowały się w różnych chromosomach, a nie w jednym (w przeciwnym razie może wystąpić zjawisko „dziedziczenia powiązanego”).
Prawo podziału będzie przestrzegane, jeśli
w hybrydach czynniki dziedziczne pozostają niezmienione;
Prawo niezależnej dystrybucji genów u potomstwa i powstawanie różnych kombinacji tych genów podczas krzyżowania dihybrydowego jest możliwe tylko w przypadku.
jeśli pary genów allelicznych znajdują się w różnych parach homologicznych chromosomów.
Naruszenie tych warunków może prowadzić albo do braku rozszczepienia w drugim pokoleniu, albo do rozszczepienia w pierwszym pokoleniu; lub do zniekształcenia stosunku różnych genotypów i fenotypów. Prawa Mendla są uniwersalne dla wszystkich organizmów diploidalnych rozmnażających się płciowo. Na ogół są one ważne dla genów autosomalnych z pełną penetracją (100% częstość manifestacji analizowanej cechy; 100% penetracja oznacza, że cecha jest wyrażana u wszystkich nosicieli allelu warunkującego rozwój tej cechy) i stałą ekspresją; stała ekspresja oznacza, że nasilenie fenotypowe cechy jest takie samo lub w przybliżeniu takie samo u wszystkich nosicieli allelu determinującego rozwój tej cechy.
Wniosek.
Prawa Gregora Mendla są obecnie szeroko stosowane w hodowli roślin, zwierząt i mikroorganizmów, w medycynie, inżynierii genetycznej i wielu innych gałęziach ludzkiego życia, a także w rozwiązywaniu problemów genetyki.
Należy zauważyć, że Mendel formułował prawa i wyciągał wnioski w czasie, gdy nic nie było wiadomo o DNA, genach i chromosomach. Okazało się jednak, że miał całkowitą rację i choć nie od razu, jego teorie zostały uznane i wzięte za podstawę rozwijającej się nauki genetyki.
Mendlowska teoria dziedziczności, tj. Całość poglądów na temat uwarunkowań dziedzicznych i charakteru ich przekazywania z rodziców na potomstwo jest w swoim znaczeniu wprost przeciwstawna teoriom Domainelea, w szczególności teorii pangenezy proponowanej przez Darwina. Zgodnie z tą teorią znaki rodziców są bezpośrednie, tj. ze wszystkich części ciała, przekazywany potomstwu. Dlatego charakter cechy potomka powinien bezpośrednio zależeć od właściwości rodzica. Jest to całkowicie sprzeczne z wnioskami wyciągniętymi przez Mendla: wyznacznikami dziedziczności, tj. geny są obecne w organizmie stosunkowo niezależnie od siebie. Charakter cech (fenotyp) determinuje ich losowa kombinacja. Nie są modyfikowane przez żadną część ciała i pozostają w relacji dominacja-recesja. Mendlowska teoria dziedziczności sprzeciwia się więc idei dziedziczenia cech nabytych w trakcie indywidualnego rozwoju.
Eksperymenty Mendla posłużyły jako podstawa do rozwoju współczesnej genetyki - nauki, która bada dwie główne właściwości organizmu - dziedziczność i zmienność. Udało mu się zidentyfikować wzorce dziedziczenia dzięki całkowicie nowym podejściom metodologicznym:
1) Mendel z powodzeniem wybrał przedmiot badań;
2) przeanalizował dziedziczenie cech indywidualnych u potomstwa krzyżowanych roślin różniących się jedną, dwiema i trzema parami kontrastujących cech alternatywnych. W każdym pokoleniu prowadzono ewidencję osobno dla każdej pary tych cech;
3) nie tylko rejestrował uzyskane wyniki, ale także dokonywał ich matematycznej obróbki.
Wymienione proste metody badawcze stanowiły zasadniczo nową, hybrydologiczną metodę badania dziedziczenia, która stała się podstawą dalszych badań genetycznych.
Bibliografia.
Biologia ogólna: Podręcznik dla klas 9-10. Poślubić szkoła / Polyansky Yu.I., Brown AD, Verzilin N.M. i inni; M .: Edukacja, 1987.-287 s.: chory.
Mendla doszedł do wniosku, że predyspozycje do porodu… wpływają na częstość występowania genotypów w populacji. Prawa Mendla nie mów nam nic o częstotliwościach...Pierwsze prawo Mendla
Nazywa się krzyżowanie dwóch organizmów hybrydyzacja, potomstwo ze skrzyżowania dwóch osobników o różnej dziedziczności nazywa się hybrydowy, i osobna osoba - hybryda. Mononukleoza hybrydowy nazywa się krzyżowaniem dwóch organizmów, które różnią się od siebie jedną parą alternatywnych (wzajemnie wykluczających się) cech. W związku z tym przy takim krzyżowaniu śledzone są wzorce dziedziczenia tylko dwóch cech, których rozwój wynika z pary genów allelicznych. Wszystkie inne znaki charakterystyczne dla tych organizmów nie są brane pod uwagę.
Jeśli krzyżujesz rośliny grochu z żółtymi i zielonymi nasionami, wszystkie powstałe hybrydy będą miały żółte nasiona. Ten sam obraz obserwuje się podczas krzyżowania roślin o gładkim i pomarszczonym kształcie nasion; całe potomstwo pierwszego pokolenia będzie miało gładki kształt nasion:
Dlatego nazwano pierwsze prawo Mendla Prawo jednorodności hybryd pierwszej generacji.
Jeśli użyjemy terminów, które pojawiły się wiele lat po pracy Mendla, to możemy powiedzieć, że komórki roślin grochu jednej odmiany zawierają dwa geny tylko koloru żółtego, a geny roślin innej odmiany - dwa geny tylko koloru zielonego. Geny odpowiedzialne za rozwój jednej cechy (na przykład koloru nasion) nazywane są geny alleliczne... W konsekwencji w hybrydzie pierwszego pokolenia rozwija się tylko jedna z każdej pary cech alternatywnych. Drugi objaw wydaje się zanikać, nie pojawia się. Zjawisko dominacji w hybrydzie z znak jednego z rodziców G. Mendla nazwany domin racjonowanie. Nazwano cechę, która przejawia się w hybrydzie pierwszego pokolenia i hamuje rozwój innej cechy dominujący, i odwrotnie, czyli stłumiony, znak - recesywny. Gregor Mendel postanowił zastąpić opis cech roślin abstrakcyjnym kodem „A, B, C, D, E, F, G” oraz „a, b, c, d, e, f, g”, a następnie Los jednej pary cech przeniósł się do oglądania dwóch, trzech, czterech par jednocześnie. Duże A, B, C, D, E, F, G zaznaczył cechy dominujące; małe a, b, c, d, e, f, g - recesywne. Jeśli w genotypie organizmu (zygoty) występują dwa identyczne geny alleliczne, oba są dominujące lub oba są recesywne (AA lub aa), taki organizm nazywa się homozygotyczny. Jeśli z pary genów allelicznych jeden jest dominujący, a drugi recesywny (Aa), wtedy taki organizm nazywa się heterozygotyczny.
Prawo podziału, czyli drugie prawo Mendla.
Jeżeli potomkowie pierwszego pokolenia, identyczni w badanej cesze, zostaną ze sobą skrzyżowani, to w drugim pokoleniu cechy obojga rodziców pojawiają się w określonej proporcji liczbowej: 3/4 osobników będzie miało cechę dominującą, ¼ recesywny:

Zjawisko, w którym krzyżowanie heterozygotycznych osobniki prowadzą do powstania potomstwa, z których część rogo nosi cechę dominującą, a część - recesywny ny, nazywa się dzieleniem. W konsekwencji cecha recesywna u mieszańców pierwszego pokolenia nie zniknęła, a jedynie została stłumiona i ujawni się w drugim pokoleniu mieszańców.
Hipoteza czystości gamet. Mendel zasugerował, że podczas formowania się mieszańców czynniki dziedziczne nie mieszają się, ale pozostają niezmienione. W hybrydzie występują oba czynniki – dominujący i recesywny, przy czym dominujący czynnik dziedziczny przejawia się w postaci cechy, natomiast czynnik recesywny jest stłumiony. Komunikacja między pokoleniami podczas rozmnażania płciowego odbywa się za pośrednictwem komórek płciowych - mam meta. Dlatego należy założyć, że każda gameta zawiera tylko jeden czynnik z pary. Następnie, podczas zapłodnienia, fuzja dwóch gamet, z których każda niesie recesywny czynnik dziedziczny, doprowadzi do powstania organizmu z cechą recesywną, która objawia się fenotypowo. Fuzja gamet, z których każda zawiera czynnik dominujący, lub dwie gamety, z których jedna zawiera czynnik dominujący, a druga czynnik recesywny, prowadzi do rozwoju organizmu z cechą dominującą.
Mendel tłumaczył podział potomstwa podczas krzyżowania osobników heterozygotycznych faktem, że gamety są czyste genetycznie, to znaczy niosą tylko jeden gen z pary allelicznej. Hipoteza(teraz nazywany prawem) czystość gamety można sformułować w następujący sposób: podczas tworzenia komórek zarodkowych do każdej gamety dostaje się tylko jeden gen z pary alleli.
Dlaczego i jak to się dzieje? Wiadomo, że każda komórka ciała ma dokładnie taki sam diploidalny zestaw chromosomów. Dwa homologiczne chromosomy zawierają dwa identyczne geny. Genetycznie „czyste” gamety powstają w następujący sposób: gdy łączą się gamety męskie i żeńskie, uzyskuje się hybrydę z diploidalnym (podwójnym) zestawem chromosomów.
Jak widać na diagramie (Załącznik 2), zygota otrzymuje połowę chromosomów od organizmu ojcowskiego, a połowę od matki.
W procesie tworzenia gamet w hybrydzie chromosomy homologiczne podczas pierwszego podziału mejotycznego również wchodzą do różnych komórek.
W przypadku tej pary allelicznej powstają dwie odmiany gamet. Podczas zapłodnienia gamety niosące te same lub różne allele przypadkowo spotykają się ze sobą. Ze względu na prawdopodobieństwo statystyczne, przy odpowiednio dużej liczbie gamet u potomstwa, 25% genotypów będzie homozygotycznych dominujących, 50% - heterozygotycznych, 25% - homozygotycznych recesywnych, czyli stosunek 1AA: 2Aa: 1AA.
Odpowiednio, zgodnie z fenotypem, potomstwo drugiego pokolenia podczas krzyżowania monohybrydowego rozkłada się w proporcji 3:1 (¾ osobników z cechą dominującą, ¼ osobników z cechą recesywną).
Podstawą cytologiczną rozszczepiania cech w krzyżowaniu monohybrydowym jest dywergencja chromosomów homologicznych do różnych biegunów komórki i powstawanie haploidalnych komórek płciowych w mejozie.
W omówionych powyżej przykładach zasada jednorodności wyrażała się w tym, że wszystkie mieszańce były zewnętrznie podobne do jednego z rodziców. Nie zawsze tak jest. Często znaki w formach heterozygotycznych mają charakter pośredni, tj. dominacja może nie być pełna. Schemat skrzyżowania dwóch dziedzicznych form rośliny Night Beauty:


Jedna z nich ma czerwone kwiaty (i to jest cecha dominująca), a druga jest biała. Diagram pokazuje, że wszystkie hybrydy pierwszej generacji mają różowe kwiaty. W drugiej generacji rozszczepienie następuje w stosunku 1: 2: 1, tj. jeden czerwony kwiat (homozygota), dwa różowe kwiaty (heterozygota), jeden biały (homozygota). Zjawisko to nazywa się niepełną dominacją.
W przypadku niepełnej dominacji, dominujący gen w stanie heterozygotycznym nie zawsze całkowicie tłumi gen recesywny. W niektórych przypadkach hybryda fi nie odtwarza w pełni żadnej z cech rodzicielskich, a cecha ta ma charakter pośredni, z mniejszym lub większym odchyleniem w kierunku stanu dominującego lub recesywnego. Ale wszystkie osobniki tego pokolenia są jednolite w tej charakterystyce. Niepełna dominacja jest powszechna. Stwierdzono to w badaniach nad dziedziczeniem koloru kwiatów u lwiej paszczy, koloru wełny u bydła i owiec, cech biochemicznych u ludzi itp. Cechy pośrednie wynikające z niepełnej dominacji często reprezentują wartość estetyczną lub materialną dla człowieka. Powstaje pytanie: czy można rozmnażać się przez selekcję, na przykład odmianę nocnego piękna o różowym kolorze kwiatów? Oczywiście nie, ponieważ ta cecha rozwija się tylko u heterozygot i przy ich krzyżowaniu zawsze następuje rozszczepienie:
Niepełna dominacja jest powszechna. Stwierdzono to w badaniach nad dziedziczeniem koloru kwiatów u lwiej paszczy, koloru wełny u bydła i owiec, cech biochemicznych u ludzi itp. Cechy pośrednie wynikające z niepełnej dominacji często reprezentują wartość estetyczną lub materialną dla człowieka. Powstaje pytanie: czy można rozmnażać się przez selekcję, na przykład odmianę nocnego piękna o różowym kolorze kwiatów? Oczywiście, że nie, ponieważ ta cecha rozwija się tylko u heterozygot i przy ich krzyżowaniu zawsze następuje rozszczepienie.
Prawo niezależnej kombinacji, czyli trzecie Prawo Mendla.
Badanie Mendla nad dziedziczeniem jednej pary alleli pozwoliło ustalić szereg ważnych wzorców genetycznych: zjawisko dominacji, niezmienność alleli recesywnych w mieszańcach, podział potomstwa mieszańców w stosunku 3:1, a także założyć, że gamety są genetycznie czyste, tj. zawierają tylko jeden gen z par allelicznych. Jednak organizmy różnią się wieloma genami. Możliwe jest ustalenie wzorców dziedziczenia dwóch par cech alternatywnych i więcej poprzez dwuhybrydowy lub krzyżowanie wielohybrydowe, tj. krzyżowanie form rodzicielskich różniących się dwiema parami cech.
Naukowiec z Czech Gregor Mendel (1822-1884), w trakcie swoich badań nad krzyżowaniem różnych odmian grochu, ujawnił pewne wzorce, które obecnie są powszechnie znane pod nazwą trzech praw Mendla.
Pierwsze prawo
Jeśli okazy roślin uzyskano w pierwszym pokoleniu przez krzyżowanie homozygotycznych odmian rodzicielskich (czyste linie) o różnych cechach, to potomstwo jest takie samo pod względem genotypu i fenotypu. Przez cechę w tym przypadku rozumie się dowolną wybraną jakość roślin, dzięki której można wyróżnić dwie odmiany. Pierwsze prawo Mendla jest również znane jako zasada dominacji lub prawo jednolitości dla hybryd pierwszej generacji. Cecha, która została zidentyfikowana w pierwszym pokoleniu, została nazwana dominującą, a cecha, która została stłumiona i nie ujawniła się, była recesywna.
Drugie prawo
Prawo podziału. Jej istota polega na tym, że krzyżując np. podczas samozapylenia tego samego typu mieszańców pierwszego pokolenia w następnym pokoleniu, potomstwo dzieli się według wybranej cechy (fenotypu) w stosunku 3 : 1 w przypadku całkowitej dominacji i 1: 2: 1 w przypadku niepełnej dominacji.
Trzecie prawo
Trzecie prawo Mendla dotyczy niezależnego dziedziczenia cech lub niezależnej kombinacji genów roślin. W przypadku krzyżowania okazów roślin, które różnią się między sobą dwiema lub więcej parami cech alternatywnych, materiał genetyczny i cechy fenotypowe, które określają, są dziedziczone niezależnie od siebie i można je łączyć w dowolnej kombinacji.
Eksperymenty czeskiego naukowca stały się podstawą rozwoju nowoczesnej genetyki. G. Mendel był w stanie odkryć podstawowe wzorce dziedziczenia za pomocą najnowszych podejść metodologicznych, które są nadal stosowane przez wszystkich genetyków.
W jego eksperymentach wszystkie skrzyżowane okazy należały do tego samego gatunku roślin. Ważne jest, aby jednocześnie wyraźnie różniły się od siebie jedną, dwiema, kilkoma parami alternatywnych (przeciwstawnych) znaków. Cechy te były stałe, to znaczy były przekazywane z pokolenia na pokolenie podczas krzyżowania w obrębie tego samego gatunku. Konieczne było zbadanie potomstwa z każdej pary mieszańców z osobna oraz dokonanie ilościowego wyliczenia mieszańców różniących się pewnymi parami cech alternatywnych.
Te metody badawcze stały się podstawą nowej metody hybrydologicznej, której nadejściem rozpoczęła się era badań dziedziczności i zmienności.
Wzorce dziedziczenia cech podczas rozmnażania płciowego ustalił G. Mendel. Konieczne jest dokładne zrozumienie genotypu i fenotypu, alleli, homo- i heterozygotyczności, dominacji i jej rodzajów, rodzajów krzyżówek, a także opracowanie schematów.
Monohybrydowy nazywa się skrzyżowanie, w którym formy rodzicielskie różnią się od siebie jedną parą kontrastujących, alternatywnych znaków.
W związku z tym przy takim krzyżowaniu śledzone są wzorce dziedziczenia tylko dwóch wariantów cechy, których rozwój wynika z pary genów allelicznych. Przykładami krzyżówek monohybrydowych wykonanych przez G. Mendla są krzyżówki grochu z tak wyraźnie widocznymi cechami alternatywnymi jak fioletowo-białe kwiaty, żółto-zielona barwa niedojrzałych owoców (fasola), gładka i pomarszczona powierzchnia nasion, żółto-zielona barwa itp. .
Jednolitość mieszańców pierwszej generacji (pierwsze prawo Mendla) Krzyżując groch z fioletowymi (AA) i białymi (aa) kwiatami, Mendel odkrył, że wszystkie rośliny hybrydowe pierwszej generacji (F1) miały fioletowe kwiaty (ryc. 2).
Zdjęcie 2 – Schemat krzyżowania monohybrydowego
W tym samym czasie nie pojawił się biały kolor kwiatu. Podczas krzyżowania roślin, które mają gładkie i pomarszczone nasiona, hybrydy będą miały gładkie nasiona. G. Mendel odkrył również, że wszystkie hybrydy F 1 były jednolite (homogeniczne) dla każdej z siedmiu badanych cech. W konsekwencji u mieszańców pierwszego pokolenia z pary rodzicielskich cech alternatywnych pojawia się tylko jedna, a cecha drugiego rodzica wydaje się zanikać.
Znaki alternatywne są wzajemnie wykluczającymi się, kontrastującymi znakami.
Mendel nazwał przewagę cech jednego z rodziców w hybrydach F1 jako dominację, a odpowiadającą jej cechę jako dominującą. Cechy, które nie występują u mieszańców F 1 nazwał recesywnymi. Ponieważ wszystkie hybrydy pierwszej generacji są jednolite, zjawisko to nazwano pierwszymi prawami Mendla lub prawem jednorodności hybryd pierwszej generacji i regułą dominacji.
Można to sformułować w następujący sposób: krzyżując dwa organizmy należące do różnych czystych linii (dwa organizmy homozygotyczne), różniące się między sobą jedną parą alternatywnych cech, całe pierwsze pokolenie mieszańców będzie jednorodne i będzie nosiło cechę jednego z rodzice.
Każdy gen ma dwa stany – „A” i „a”, więc tworzą jedną parę, a każdy członek pary nazywany jest allelem. Geny zlokalizowane w tych samych loci (sekcjach) chromosomów homologicznych i determinujące alternatywny rozwój tej samej cechy nazywane są allelicznymi.
Na przykład fioletowy i biały kolor kwiatu grochu jest odpowiednio dominujący i recesywny dla dwóch alleli (A i a) jednego genu. Ze względu na obecność dwóch alleli możliwe są dwa stany organizmu: homo- i heterozygotyczny. Jeśli organizm zawiera te same allele danego genu (AA lub aa), to nazywa się go homozygotą dla tego genu (lub cechy), a jeśli inny (Aa), to heterozygotą. Stąd allel jest formą istnienia genu. Przykładem genu triallelicznego jest gen determinujący układ grup krwi ABO u ludzi. Alleli jest więcej: w przypadku genu kontrolującego syntezę ludzkiej hemoglobiny są ich dziesiątki.
Z hybrydowych nasion grochu Mendel wyhodował rośliny, które poddał samozapyleniu, a powstałe nasiona ponownie zasiał. W rezultacie powstała druga generacja hybryd, czyli hybrydy F2. Wśród tych ostatnich stwierdzono rozszczepienie dla każdej pary cech alternatywnych w stosunku około 3:1, tj. trzy czwarte roślin miało cechy dominujące (fioletowe kwiaty, żółte nasiona, gładkie nasiona itp.), a jedna czwarta była recesywne (białe kwiaty, zielone nasiona, pomarszczone nasiona itp.). W konsekwencji cecha recesywna w hybrydzie F1 nie zniknęła, a jedynie została stłumiona i pojawiła się ponownie w drugim pokoleniu. To uogólnienie nazwano później drugim prawem Mendla, czyli prawem rozszczepienia.
Rozszczepienie to zjawisko, w którym krzyżowanie osobników heterozygotycznych prowadzi do powstania potomstwa, z których część jest dominująca, a część recesywna.
Drugie prawo Mendla: gdy krzyżuje się ze sobą dwóch potomków pierwszego pokolenia (dwa osobniki heterozygotyczne) w drugim pokoleniu obserwuje się rozszczepienie w pewnym stosunku liczbowym: zgodnie z fenotypem 3:1, zgodnie z genotypem 1:2 : 1 (rys. 3).

Rysunek 3 - Schemat znaków podziału
przy krzyżowaniu mieszańców F 1
Podział cech u potomstwa podczas krzyżowania osobników heterozygotycznych został wyjaśniony przez G. Mendla faktem, że gamety są czyste genetycznie, to znaczy niosą tylko jeden gen z pary allelicznej. Prawo czystości gamet można sformułować w następujący sposób: podczas tworzenia komórek zarodkowych do każdej gamety dostaje się tylko jeden gen z pary alleli.
Należy pamiętać, że zastosowanie metody hybrydologicznej do analizy dziedziczenia cech u dowolnego gatunku zwierząt lub roślin obejmuje następujące krzyżówki:
krzyżowanie form rodzicielskich (P), różniących się jedną (krzyżowanie monohybrydowe) lub kilkoma parami (krzyżowanie polihybrydowe) cech alternatywnych i uzyskaniem mieszańców pierwszego pokolenia (F 1);
krzyżowanie mieszańców F 1 ze sobą i uzyskanie mieszańców drugiej generacji (F 2);
analiza matematyczna wyników krzyżowania.
Następnie Mendel zajął się badaniem krzyżowania dihybrydów.
Krzyżówka dihybrydowa Jest krzyżówką, w której zaangażowane są dwie pary alleli (sparowane geny są allelami i znajdują się tylko w chromosomach homologicznych).
Podczas krzyżowania dihybrydów G. Mendel badał dziedziczenie cech, które odpowiadają za geny leżące w różnych parach homologicznych chromosomów. W związku z tym każda gameta musi zawierać jeden gen z każdej pary allelicznej.
Hybrydy heterozygotyczne dla dwóch genów nazywane są diheterozygotycznymi, a jeśli różnią się trzema lub wieloma genami, nazywane są odpowiednio tri- i poliheterozygotycznymi.
Bardziej złożone schematy krzyżówek dihybrydowych, rejestrację genotypów i fenotypów F 2 przeprowadza się przy użyciu siatki Pennetta. Rozważ przykład takiego krzyża. Do krzyżowania pobrano dwie początkowe homozygotyczne formy rodzicielskie: pierwsza forma miała żółte i gładkie nasiona; druga forma miała zielone i pomarszczone nasiona (ryc. 4).

Rysunek 4 - Krzyżowanie dihybrydowe roślin grochu,
inny kształt i kolor nasion
Dominują żółte zabarwienie i gładkie nasiona; zielony kolor i pomarszczone nasiona są objawami recesywnymi. Hybrydy pierwszej generacji - skrzyżowane ze sobą. W drugim pokoleniu zaobserwowano rozszczepienie fenotypowe w stosunku 9:3:3:1 lub (3+1) 2, po samozapyleniu mieszańców F1 zgodnie z prawem rozszczepiania, ponownie pojawiły się pomarszczone i zielone nasiona.
Rośliny rodzicielskie mają w tym przypadku genotypy AABB i aabb, a genotyp mieszańców F1 - AaBb jest diheterozygotyczny.
Zatem w przypadku krzyżowania osobników heterozygotycznych różniących się kilkoma parami cech alternatywnych u potomstwa obserwuje się rozszczepienie fenotypowe w stosunku (3 + 1) n, gdzie n jest liczbą par cech alternatywnych.
Geny, które determinują rozwój różnych par cech, nazywane są nie allelicznymi.
Wyniki krzyżowania dihybrydowego i polihybrydowego zależą od tego, czy geny determinujące rozważane cechy znajdują się w tych samych, czy w różnych chromosomach. Mendel natrafił na cechy, których geny znajdowały się w różnych parach homologicznych chromosomów grochu.
W mejozie homologiczne chromosomy różnych par są losowo łączone w gamety. Jeśli chromosom ojcowski z pierwszej pary dostanie się do gamety, to z równym prawdopodobieństwem do tej gamety może dostać się zarówno ojcowski, jak i matczyny chromosom drugiej pary. Dlatego cechy, których geny znajdują się w różnych parach homologicznych chromosomów, są łączone niezależnie od siebie. Następnie okazało się, że z siedmiu par cech badanych przez Mendla grochu, w których diploidalna liczba chromosomów wynosi 2 n = 14, geny odpowiedzialne za jedną z par cech znajdowały się na tym samym chromosomie. Mendel nie stwierdził jednak naruszenia prawa niezależnego dziedziczenia, ponieważ nie zaobserwowano powiązania między tymi genami ze względu na dużą odległość między nimi).
Na podstawie swoich badań Mendel wydedukował trzecie prawo – prawo niezależnego dziedziczenia cech, czyli niezależnego łączenia genów.
Każda para genów allelicznych (i kontrolowanych przez nie alternatywnych cech) jest dziedziczona niezależnie od siebie.
Prawo niezależnego łączenia genów stanowi podstawę kombinacyjnej zmienności obserwowanej podczas krzyżowania we wszystkich żywych organizmach. Zauważmy również, że w przeciwieństwie do pierwszego prawa Mendla, które zawsze jest prawdziwe, drugie prawo jest ważne tylko dla genów zlokalizowanych w różnych parach homologicznych chromosomów. Wynika to z faktu, że niehomologiczne chromosomy są łączone w komórce niezależnie od siebie, co zostało udowodnione nie tylko poprzez badanie natury dziedziczenia cech, ale także bezpośrednią metodą cytologiczną.
Studiując materiał, zwróć uwagę na przypadki naruszeń regularnego rozszczepiania przez fenotyp spowodowany śmiertelnym działaniem poszczególnych genów.
Dziedziczność i zmienność. Dziedziczność i zmienność to najważniejsze właściwości charakterystyczne dla wszystkich żywych organizmów.
Zmienność dziedziczna lub genotypowa dzieli się na kombinacyjną i mutacyjną.
Zmienność nazywa się kombinacyjną, która polega na tworzeniu rekombinacji, czyli takich kombinacji genów, których rodzice nie mieli.
U podstaw kombinacyjnej zmienności leży rozmnażanie płciowe organizmów, w wyniku którego powstaje ogromna różnorodność genotypów. Trzy procesy są praktycznie nieograniczonymi źródłami zmienności genetycznej:
Niezależna dywergencja chromosomów homologicznych w pierwszym podziale mejotycznym. To właśnie niezależne połączenie chromosomów w mejozie jest podstawą trzeciego prawa G. Mendla. Pojawienie się zielonych, gładkich i żółto pomarszczonych nasion grochu w drugim pokoleniu ze skrzyżowania roślin o gładkich żółto-zielono pomarszczonych nasionach jest przykładem kombinacyjnej zmienności.
Wzajemna wymiana regionów chromosomów homologicznych lub krzyżowanie. Tworzy nowe grupy sprzężenia, czyli służy jako ważne źródło rekombinacji genetycznej alleli. W zygocie zrekombinowane chromosomy przyczyniają się do pojawienia się cech nietypowych dla każdego z rodziców.
Losowe połączenie gamet podczas zapłodnienia.
Te źródła kombinatywnej zmienności działają niezależnie i jednocześnie, zapewniając przy tym stałe „przetasowanie” genów, co prowadzi do pojawienia się organizmów o innym genotypie i fenotypie (same geny się nie zmieniają). Jednak nowe kombinacje genów dość łatwo ulegają degradacji, gdy są przekazywane z pokolenia na pokolenie.
Przykład zmienności kombinacyjnej. Kwiat urody nocy ma gen dla czerwonego koloru płatków A i gen dla białego a. Ciało Aa ma różowe płatki. Tak więc piękność nocy nie ma genu różu, różowy pojawia się, gdy połączenie (kombinacja) genu czerwonego i białego.
Osoba ma chorobę dziedziczną anemię sierpowatą. AA to norma, aa to śmierć, Aa to SKA. W przypadku SKA osoba nie może tolerować zwiększonego wysiłku fizycznego, podczas gdy nie cierpi na malarię, to znaczy czynnik wywołujący malarię Malaria Plasmodium nie może żywić się niewłaściwą hemoglobiną. Ta funkcja jest przydatna w pasie równikowym; nie ma na to genu, powstaje z kombinacji genów A i a.
W związku z tym zmienność dziedziczna jest wzmocniona przez zmienność kombinacyjną. Po powstaniu poszczególne mutacje znajdują się w sąsiedztwie innych mutacji, są częścią nowych genotypów, czyli powstaje wiele kombinacji alleli. Każda osoba jest genetycznie unikalna (z wyjątkiem bliźniąt jednojajowych i osobników, które powstały w wyniku bezpłciowego rozmnażania klonu z pojedynczą komórką jako przodkiem). Jeśli więc założymy, że w każdej parze chromosomów homologicznych jest tylko jedna para genów allelicznych, to dla osoby, której zestaw haploidalny chromosomów wynosi 23, liczba możliwych genotypów będzie wynosić od 3 do 23 stopni. Tak ogromna liczba genotypów jest 20 razy większa od liczby wszystkich ludzi na Ziemi. Jednak w rzeczywistości chromosomy homologiczne różnią się kilkoma genami i obliczenia nie uwzględniają zjawiska krzyżowania. . Dlatego liczba możliwych genotypów jest wyrażona w liczbach astronomicznych i można śmiało stwierdzić, że występowanie dwóch identycznych osób jest praktycznie nieprawdopodobne (z wyjątkiem identycznych bliźniąt, które powstały z tego samego zapłodnionego jaja). Wynika stąd w szczególności możliwość wiarygodnej identyfikacji osoby na podstawie pozostałości żywych tkanek, potwierdzenia lub wykluczenia ojcostwa.
Tak więc wymiana genów spowodowana krzyżowaniem chromosomów w pierwszym podziale mejozy, niezależna i losowa rekombinacja chromosomów w mejozie oraz szansa fuzji gamet w procesie płciowym to trzy czynniki, które zapewniają istnienie kombinacyjnej zmienności. Zmienność mutacyjna samego genotypu.
Mutacje to nagłe, dziedziczne zmiany w materiale genetycznym, prowadzące do zmiany pewnych cech organizmu.
Główne założenia teorii mutacji zostały opracowane przez naukowca G. De Vriesa w 1901 roku – 1903 i sprowadza się do tego:
Mutacje pojawiają się nagle, skokowo, jako dyskretne zmiany cech;
W przeciwieństwie do zmian niedziedzicznych, mutacje są zmianami jakościowymi przekazywanymi z pokolenia na pokolenie;
Mutacje manifestują się na różne sposoby i mogą być zarówno korzystne, jak i szkodliwe, dominujące i recesywne;
Prawdopodobieństwo wykrycia mutacji zależy od liczby przebadanych osobników;
Podobne mutacje mogą się powtarzać;
Mutacje są bezkierunkowe (spontaniczne), co oznacza, że każda część chromosomu może ulec mutacji, powodując zmiany zarówno w drobnych, jak i życiowych objawach.
Prawie każda zmiana w strukturze lub liczbie chromosomów, w których komórka zachowuje zdolność do samoreprodukowania, powoduje dziedziczną zmianę cech organizmu.
Ze względu na charakter zmiany w genomie, to znaczy zestaw genów zawartych w haploidalnym zestawie chromosomów, rozróżnia się mutacje genów, chromosomów i genomów.
Mutacje genowe lub punktowe są wynikiem zmiany sekwencji nukleotydowej w cząsteczce DNA w obrębie jednego genu.
Taka zmiana w genie jest odtwarzana podczas transkrypcji w strukturze i-RNA; prowadzi do zmiany sekwencji aminokwasów w łańcuchu polipeptydowym powstającym podczas translacji na rybosomach. W rezultacie syntetyzowane jest kolejne białko, co prowadzi do zmiany odpowiedniej cechy organizmu. Jest to najczęstszy rodzaj mutacji i najważniejsze źródło dziedzicznej zmienności organizmów.
Mutacje chromosomowe (rearanżacje lub aberracje) to zmiany w strukturze chromosomów, które można zidentyfikować i zbadać pod mikroskopem świetlnym.
Znane są rearanżacje różnych typów:
Brak – utrata końców chromosomu;
Usunięcie – utrata części chromosomu w jego środkowej części;
Powielanie – dwu- lub wielokrotne powtórzenie genów zlokalizowanych w określonej części chromosomu;
Inwersja – obrót sekcji chromosomu o 180 °, w wyniku czego geny w tej sekcji znajdują się w sekwencji przeciwnej do zwykłej;
Translokacja – zmiana pozycji dowolnej części chromosomu w zestawie chromosomów. Najczęstszym rodzajem translokacji jest wzajemna, w której obszary są wymieniane między dwoma niehomologicznymi chromosomami. Część chromosomu może zmienić swoją pozycję bez wzajemnej wymiany, pozostając w tym samym chromosomie lub będąc częścią innego.
Mutacje genomowe - zmiana liczby chromosomów w genomie komórek organizmu. Zjawisko to zachodzi w dwóch kierunkach: w kierunku wzrostu liczby całych zestawów haploidów (poliploidalność) oraz w kierunku utraty lub włączenia poszczególnych chromosomów (aneuploidia).
Poliploidalność – wielokrotny wzrost haploidalnego zestawu chromosomów. Komórki o różnej liczbie haploidalnych zestawów chromosomów nazywane są triploidami (3 n), tetraploidalnymi (4 n), heksaploidalnymi (6 n), oktaploidalnymi (8 n) itp. Najczęściej poliploidy powstają, gdy kolejność rozbieżności chromosomów bieguny komórek podczas mejozy są zaburzone lub mitoza. Poliploidalność prowadzi do zmiany cech organizmu i dlatego jest ważnym źródłem zmienności ewolucji i selekcji, zwłaszcza roślin. Wynika to z faktu, że hermafrodytyzm (samozapylenie), apomixis (partenogeneza) i rozmnażanie wegetatywne są bardzo rozpowszechnione w organizmach roślinnych. Dlatego około jedna trzecia gatunków roślin powszechnych na naszej planecie, – poliploidy, aw ostrych kontynentalnych warunkach wysokogórskich Pamirów rośnie do 85% poliploidów. Prawie wszystkie rośliny uprawne to także poliploidy, które w przeciwieństwie do swoich dzikich krewnych mają większe kwiaty, owoce i nasiona, a więcej składników odżywczych gromadzi się w organach magazynujących (łodyga, bulwy). Poliploidy łatwiej przystosowują się do niesprzyjających warunków życia, łatwiej tolerują niskie temperatury i suszę. Dlatego są szeroko rozpowszechnione w regionach północnych i wysokogórskich.